Department of Pharmaceutical and Medicinal Chemistry, Christian-Albrechts-University of Kiel, Kiel, Germany.
Department of Pharmaceutical and Medicinal Chemistry, Christian-Albrechts-University of Kiel, Kiel, Germany.
J Biol Chem. 2022 Jan;298(1):101513. doi: 10.1016/j.jbc.2021.101513. Epub 2021 Dec 18.
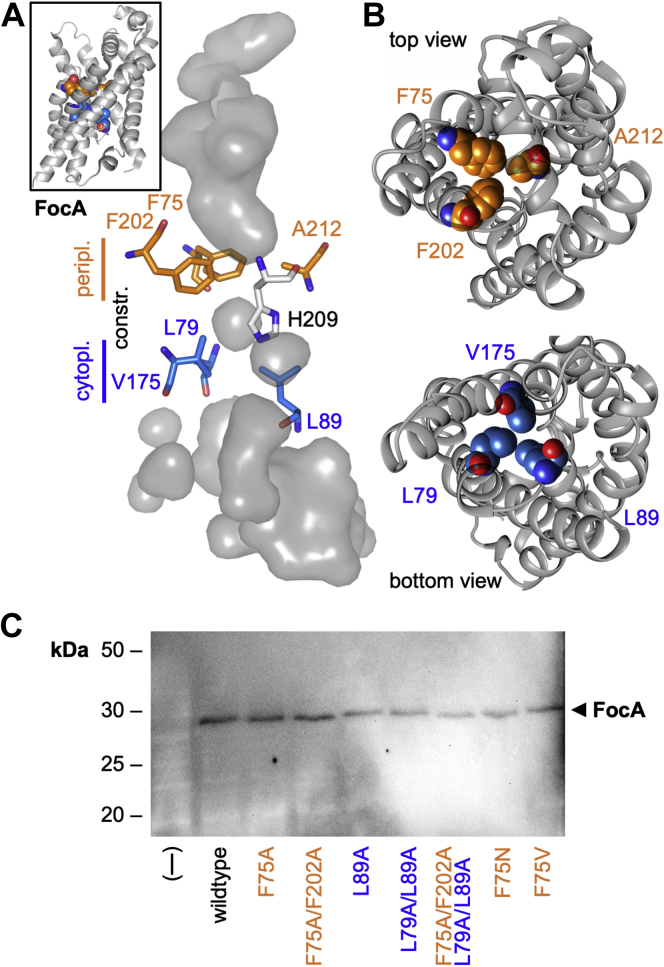
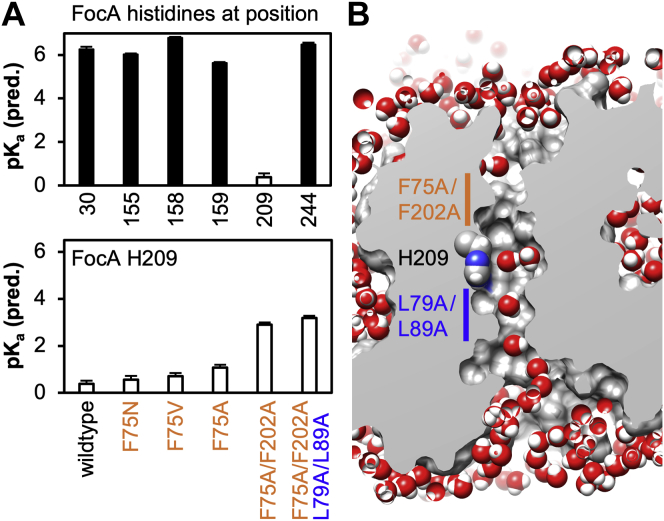
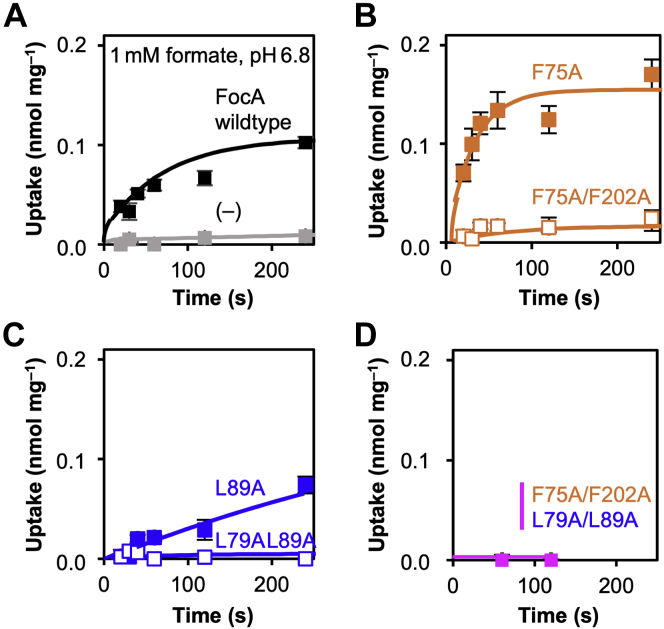
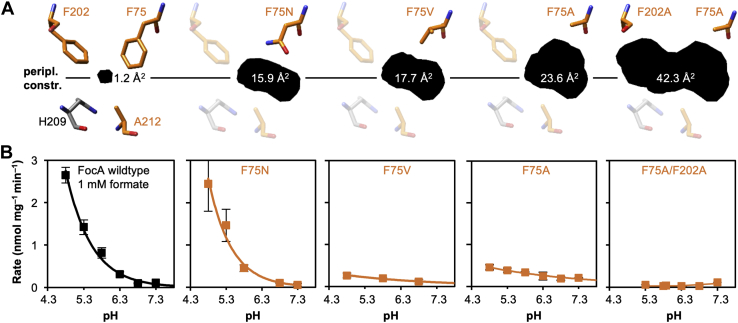
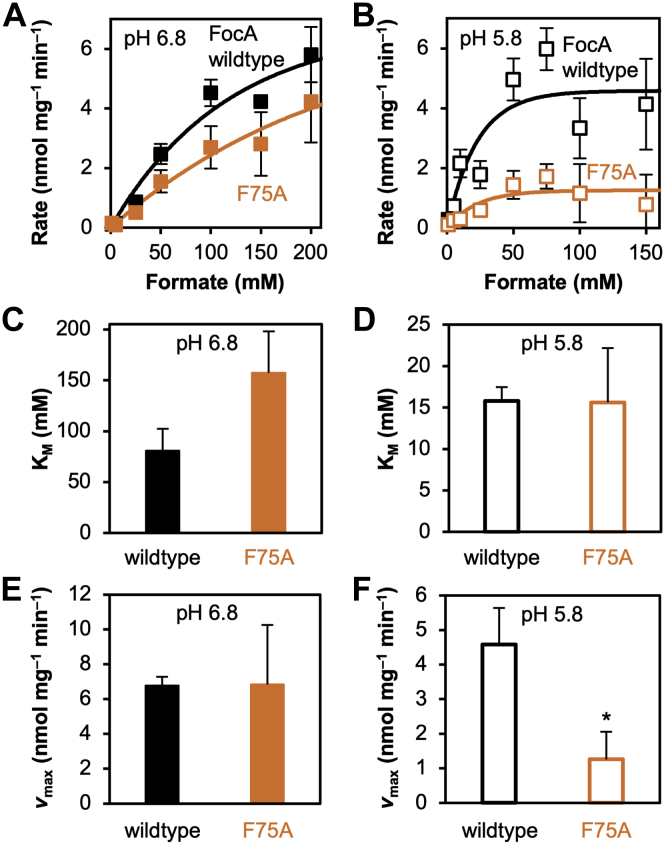
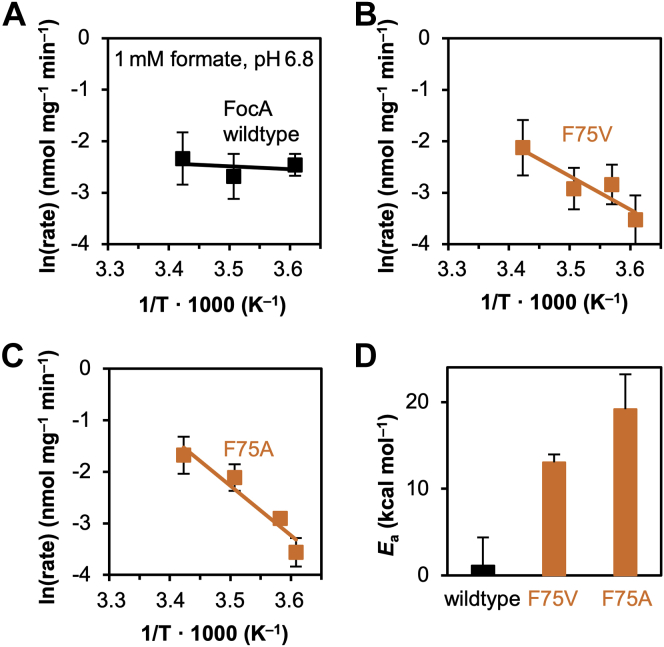
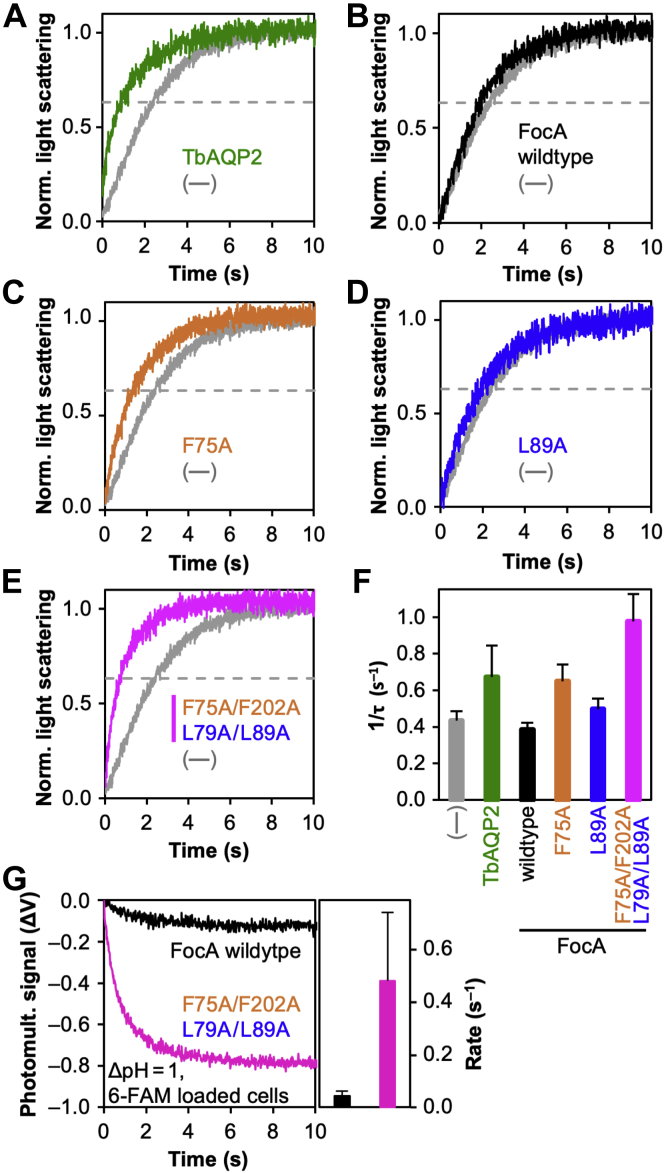
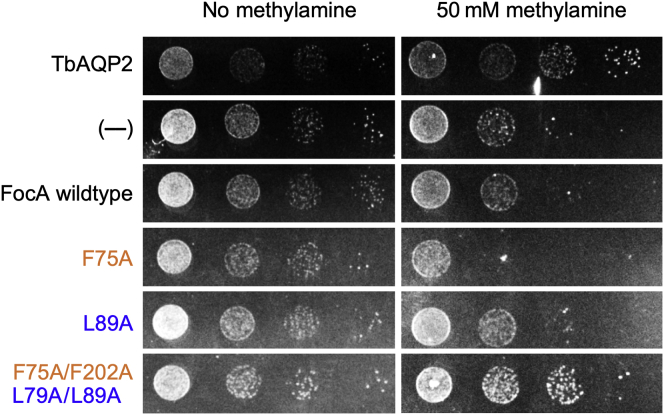
The unrelated protein families of the microbial formate-nitrite transporters (FNTs) and aquaporins (AQP) likely adapted the same protein fold through convergent evolution. FNTs facilitate weak acid anion/H cotransport, whereas AQP water channels strictly exclude charged substrates including protons. The FNT channel-like transduction pathway bears two lipophilic constriction sites that sandwich a highly conserved histidine residue. Because of lacking experiments, the function of these constrictions is unclear, and the protonation status of the central histidine during substrate transport remains a matter of debate. Here, we introduced constriction-widening mutations into the prototypical FNT from Escherichia coli, FocA, and assayed formate/H transport properties, water/solute permeability, and proton conductance. We found that enlargement of these constrictions concomitantly decreased formate/formic acid transport. In contrast to wildtype FocA, the mutants were unable to make use of a transmembrane proton gradient as a driving force. A construct in which both constrictions were eliminated exhibited water permeability, similar to AQPs, although accompanied by a proton conductance. Our data indicate that the lipophilic constrictions mainly act as barriers to isolate the central histidine from the aqueous bulk preventing protonation via proton wires. These results are supportive of an FNT transport model in which the central histidine is uncharged, and weak acid substrate anion protonation occurs in the vestibule regions of the transporter before passing the constrictions.
微生物甲酸-亚硝酸盐转运蛋白(FNTs)和水通道蛋白(AQP)的无关联蛋白家族可能通过趋同进化适应了相同的蛋白质折叠。FNTs 促进弱酸阴离子/H 共转运,而 AQP 水通道严格排除包括质子在内的带电荷底物。FNT 通道样转导途径具有两个亲脂性收缩位点,夹在高度保守的组氨酸残基之间。由于缺乏实验,这些收缩的功能尚不清楚,并且在底物转运过程中中心组氨酸的质子化状态仍然存在争议。在这里,我们在源自大肠杆菌的典型 FNT(FocA)中引入了收缩加宽突变,并检测了甲酸盐/H 转运特性、水/溶质通透性和质子电导率。我们发现这些收缩的扩大同时降低了甲酸盐/甲酸的转运。与野生型 FocA 相比,突变体无法利用跨膜质子梯度作为驱动力。消除了两个收缩的构建体表现出类似于 AQP 的水通透性,尽管伴随着质子电导率。我们的数据表明,亲脂性收缩主要作为屏障,将中央组氨酸与水相隔离,防止通过质子线进行质子化。这些结果支持 FNT 转运模型,其中中央组氨酸不带电荷,并且弱酸底物阴离子的质子化发生在转运体的前庭区域,然后通过收缩。