Department of Microbiology and Immunology, University of Oklahoma Health Sciences Centergrid.266902.9, Oklahoma City, Oklahoma, USA.
Department of Biology, Rogers State Universitygrid.440984.5, Claremore, Oklahoma, USA.
mSphere. 2022 Feb 23;7(1):e0079321. doi: 10.1128/msphere.00793-21. Epub 2022 Feb 2.
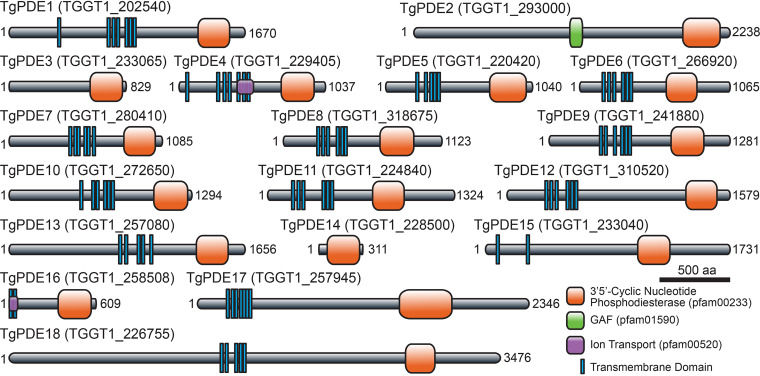
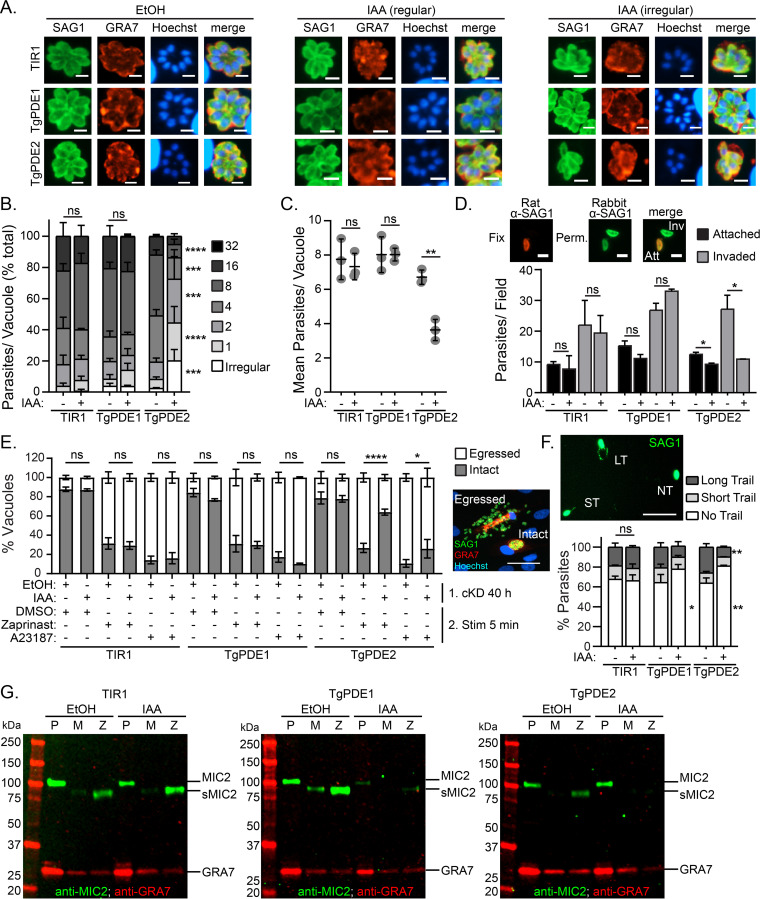
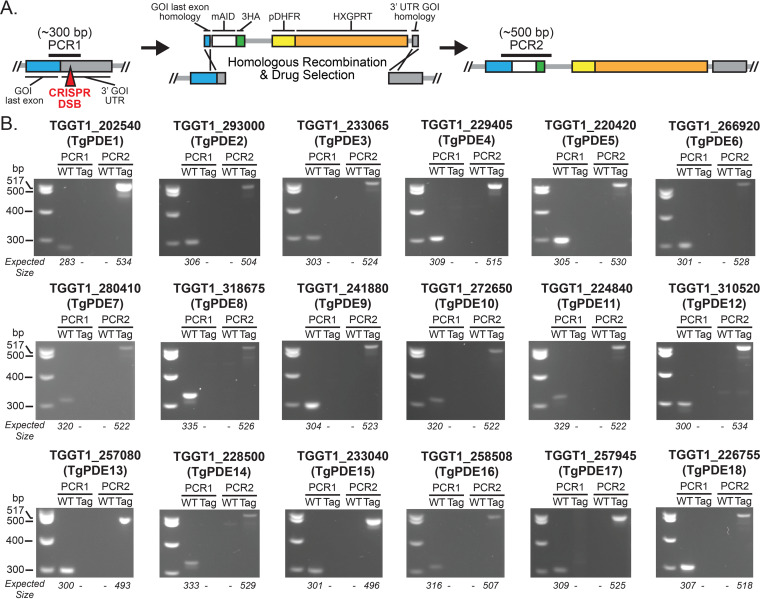
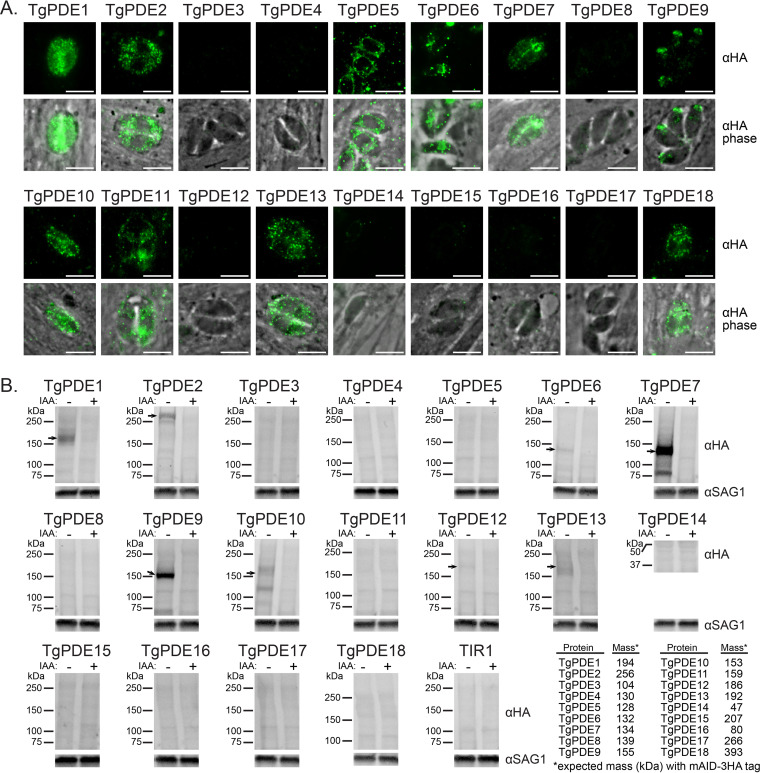
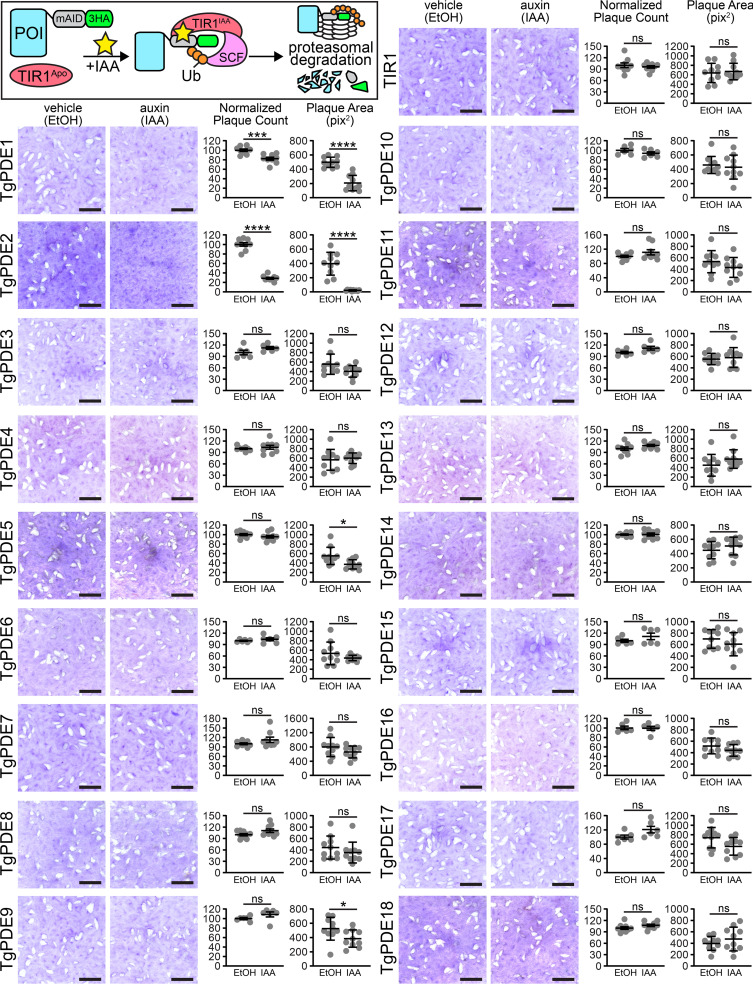
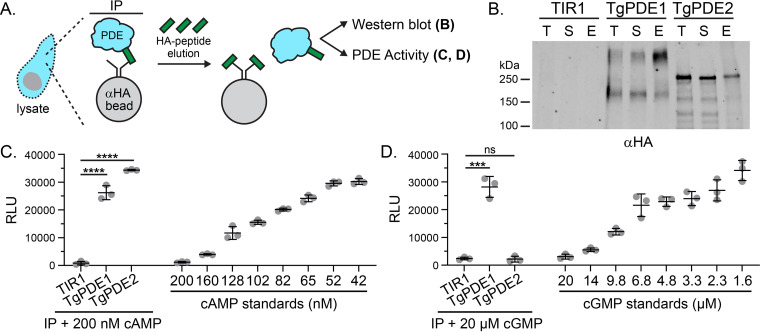
motility is both activated and suppressed by 3',5'-cyclic nucleotide signaling. Cyclic GMP (cGMP) signaling through Toxoplasma gondii protein kinase G (TgPKG) activates motility, whereas cyclic AMP (cAMP) signaling through TgPKAc1 inhibits motility. Despite their importance, it remains unclear how cGMP and cAMP levels are maintained in . Phosphodiesterases (PDEs) are known to inactivate cyclic nucleotides and are highly expanded in the genome. Here, we analyzed the expression and function of the 18-member TgPDE family in tachyzoites, the virulent life stage of . We detected the expression of 11 of 18 TgPDEs, confirming prior expression studies. A knockdown screen of the TgPDE family revealed four TgPDEs that contribute to lytic growth (TgPDE1, TgPDE2, TgPDE5, and TgPDE9). Depletion of TgPDE1 or TgPDE2 caused severe growth defects, prompting further investigation. While TgPDE1 was important for extracellular motility, TgPDE2 was important for host cell invasion, parasite replication, host cell egress, and extracellular motility. TgPDE1 displayed a plasma membrane/cytomembranous distribution, whereas TgPDE2 displayed an endoplasmic reticulum/cytomembranous distribution. Biochemical analysis of TgPDE1 and TgPDE2 purified from lysates revealed that TgPDE1 hydrolyzes both cGMP and cAMP, whereas TgPDE2 was cAMP specific. Interactome studies of TgPDE1 and TgPDE2 indicated that they do not physically interact with each other or other TgPDEs but may be regulated by kinases and proteases. Our studies have identified TgPDE1 and TgPDE2 as central regulators of tachyzoite cyclic nucleotide levels and enable future studies aimed at determining how these enzymes are regulated and cooperate to control motility and growth. Apicomplexan parasites require motility to actively infect host cells and cause disease. Cyclic nucleotide signaling governs apicomplexan motility, but it is unclear how cyclic nucleotide levels are maintained in these parasites. In search of novel regulators of cyclic nucleotides in the model apicomplexan , we identified and characterized two catalytically active phosphodiesterases, TgPDE1 and TgPDE2, that are important for s virulent tachyzoite life cycle. Enzymes that generate, sense, or degrade cyclic nucleotides make attractive targets for therapies aimed at paralyzing and killing apicomplexan parasites.
运动是由 3',5'-环核苷酸信号激活和抑制的。环鸟苷酸(cGMP)信号通过刚地弓形虫蛋白激酶 G(TgPKG)激活运动,而环腺苷酸(cAMP)信号通过 TgPKAc1 抑制运动。尽管它们很重要,但 cGMP 和 cAMP 水平如何在 中维持仍不清楚。磷酸二酯酶(PDEs)已知可使环核苷酸失活,并且在 基因组中高度扩展。在这里,我们分析了 18 个成员的 TgPDE 家族在速殖子中的表达和功能,速殖子是 的有毒生活阶段。我们检测到 18 个 TgPDE 中的 11 个表达,证实了先前的表达研究。对 TgPDE 家族的敲低筛选显示了四个对裂解生长有贡献的 TgPDE(TgPDE1、TgPDE2、TgPDE5 和 TgPDE9)。TgPDE1 或 TgPDE2 的耗竭导致严重的生长缺陷,促使进一步研究。虽然 TgPDE1 对细胞外运动很重要,但 TgPDE2 对宿主细胞入侵、寄生虫复制、宿主细胞逸出和细胞外运动很重要。TgPDE1 显示出质膜/细胞质膜的分布,而 TgPDE2 显示出内质网/细胞质膜的分布。从 裂解物中纯化的 TgPDE1 和 TgPDE2 的生化分析表明,TgPDE1 水解 cGMP 和 cAMP,而 TgPDE2 是 cAMP 特异性的。TgPDE1 和 TgPDE2 的相互作用研究表明,它们彼此之间不相互作用,也不与其他 TgPDE 相互作用,但可能受到激酶和蛋白酶的调节。我们的研究确定了 TgPDE1 和 TgPDE2 是速殖子环核苷酸水平的中央调节剂,并为未来旨在确定这些酶如何被调节以及如何合作来控制 的运动和生长的研究奠定了基础。 顶复门寄生虫需要运动才能主动感染宿主细胞并引起疾病。环核苷酸信号调节顶复门运动,但尚不清楚这些寄生虫中如何维持环核苷酸水平。在寻找模型顶复门 中新型环核苷酸调节剂的过程中,我们鉴定并表征了两种具有催化活性的磷酸二酯酶,TgPDE1 和 TgPDE2,它们对 s 的有毒速殖子生命周期很重要。产生、感知或降解环核苷酸的酶是针对瘫痪和杀死顶复门寄生虫的治疗方法的有吸引力的靶标。