Department of Cardiovascular and Metabolic Sciences, Lerner Research Institute, Cleveland Clinic, Cleveland, OH 44195.
Department of Chemistry, Cleveland State University, Cleveland, OH 44115.
Proc Natl Acad Sci U S A. 2022 Feb 15;119(7). doi: 10.1073/pnas.2115799119.
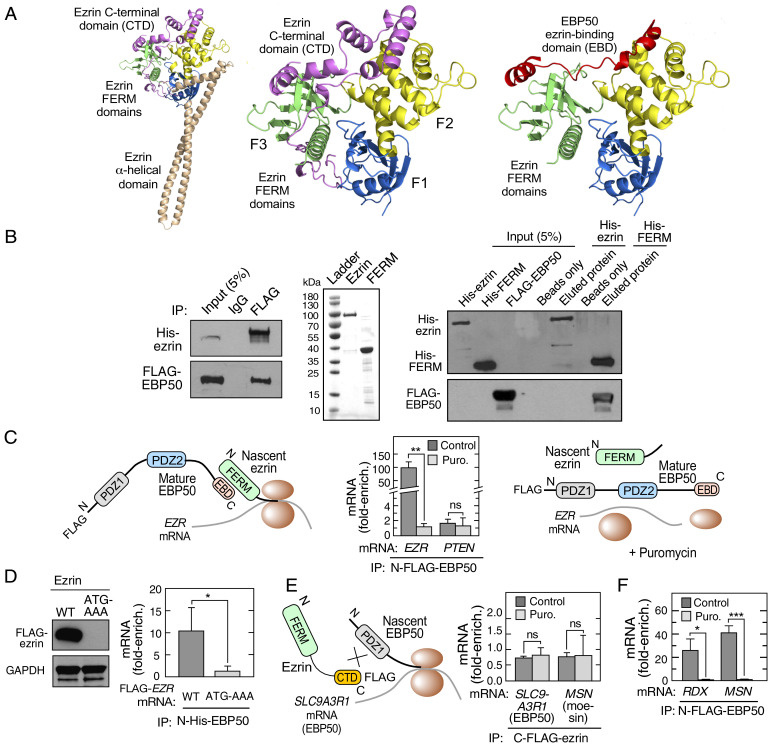
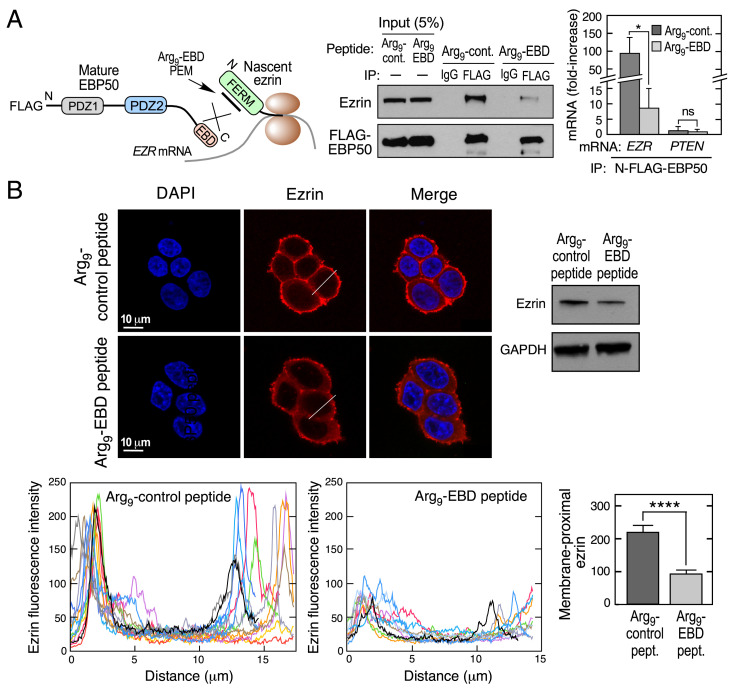
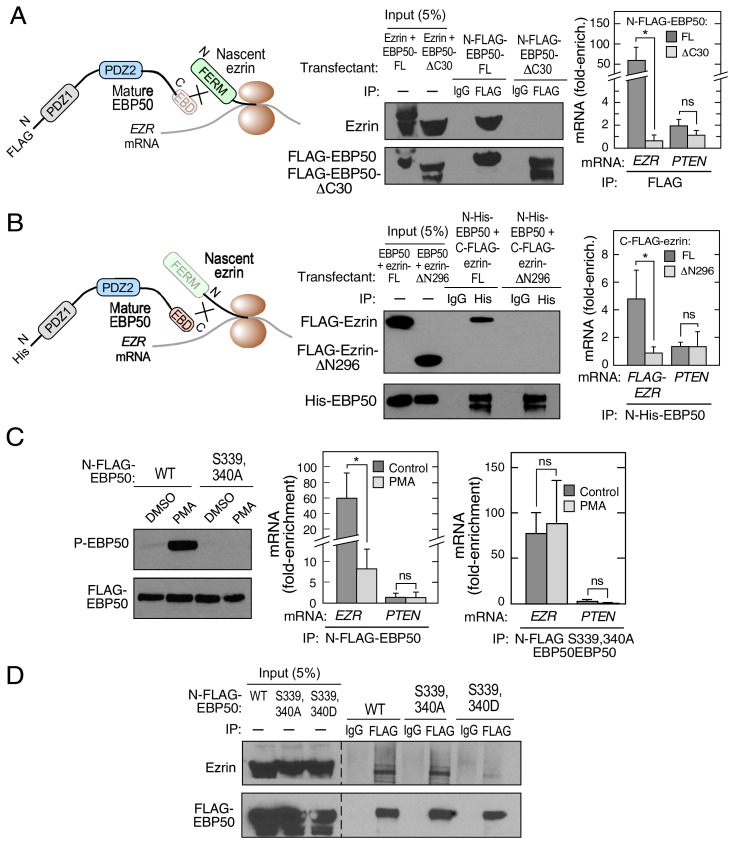
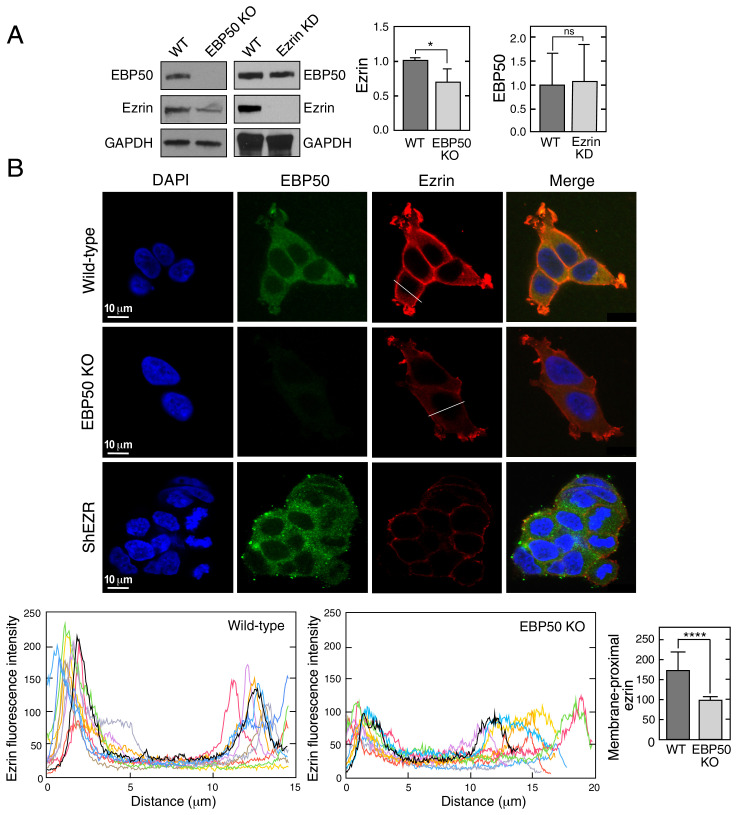
Multiprotein assemblages are the intracellular workhorses of many physiological processes. Assembly of constituents into complexes can be driven by stochastic, domain-dependent, posttranslational events in which mature, folded proteins specifically interact. However, inaccessibility of interacting surfaces in mature proteins (e.g., due to "buried" domains) can obstruct complex formation. Mechanisms by which multiprotein complex constituents overcome topological impediments remain enigmatic. For example, the heterodimeric complex formed by EBP50 and ezrin must address this issue as the EBP50-interacting domain in ezrin is obstructed by a self-interaction that occupies the EBP50 binding site. Here, we show that the EBP50-ezrin complex is formed by a cotranslational mechanism in which the C terminus of mature, fully formed EBP50 binds the emerging, ribosome-bound N-terminal FERM domain of ezrin during mRNA translation. Consistent with this observation, a C-terminal EBP50 peptide mimetic reduces the cotranslational interaction and abrogates EBP50-ezrin complex formation. Phosphorylation of EBP50 at Ser and Ser abrogates the cotranslational interaction and inhibits complex formation. In summary, we show that the function of eukaryotic mRNA translation extends beyond "simple" generation of a linear peptide chain that folds into a tertiary structure, potentially for subsequent complex assembly; importantly, translation can facilitate interactions with sterically inaccessible domains to form functional multiprotein complexes.
多蛋白组装体是许多生理过程的细胞内主力军。组成成分组装成复合物可以通过随机的、依赖结构域的、翻译后事件驱动,其中成熟的、折叠的蛋白质特异性地相互作用。然而,成熟蛋白质中相互作用表面的不可及(例如,由于“埋藏”的结构域)可能会阻碍复合物的形成。多蛋白复合物成分克服拓扑障碍的机制仍然是神秘的。例如,EBP50 和 ezrin 形成的异二聚体复合物必须解决这个问题,因为 ezrin 中的 EBP50 相互作用结构域被占据 EBP50 结合位点的自我相互作用所阻碍。在这里,我们表明 EBP50-ezrin 复合物是通过共翻译机制形成的,其中成熟的、完全形成的 EBP50 的 C 末端在 mRNA 翻译过程中与 ezrin 的核糖体结合的新兴 N 端 FERM 结构域结合。与这一观察结果一致,C 端 EBP50 肽模拟物降低了共翻译相互作用并破坏了 EBP50-ezrin 复合物的形成。EBP50 在丝氨酸和丝氨酸的磷酸化破坏了共翻译相互作用并抑制了复合物的形成。总之,我们表明真核 mRNA 翻译的功能不仅限于“简单”生成折叠成三级结构的线性肽链,为随后的复合物组装做准备;重要的是,翻译可以促进与空间不可及的结构域的相互作用,从而形成功能性的多蛋白复合物。