Department of Biophysics, Howard Hughes Medical Institute, UT Southwestern Medical Center, Dallas, TX, USA.
Department of Biological Chemistry and Molecular Pharmacology, Harvard Medical School, Boston, MA, USA.
Nat Struct Mol Biol. 2022 Apr;29(4):320-328. doi: 10.1038/s41594-022-00743-5. Epub 2022 Mar 24.
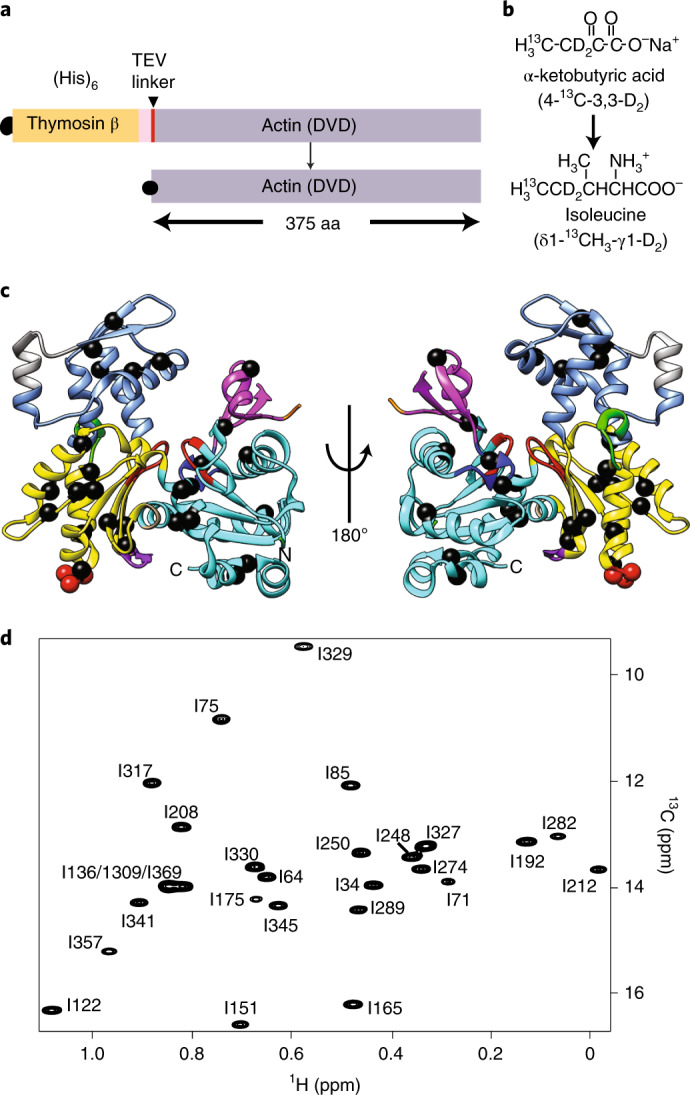
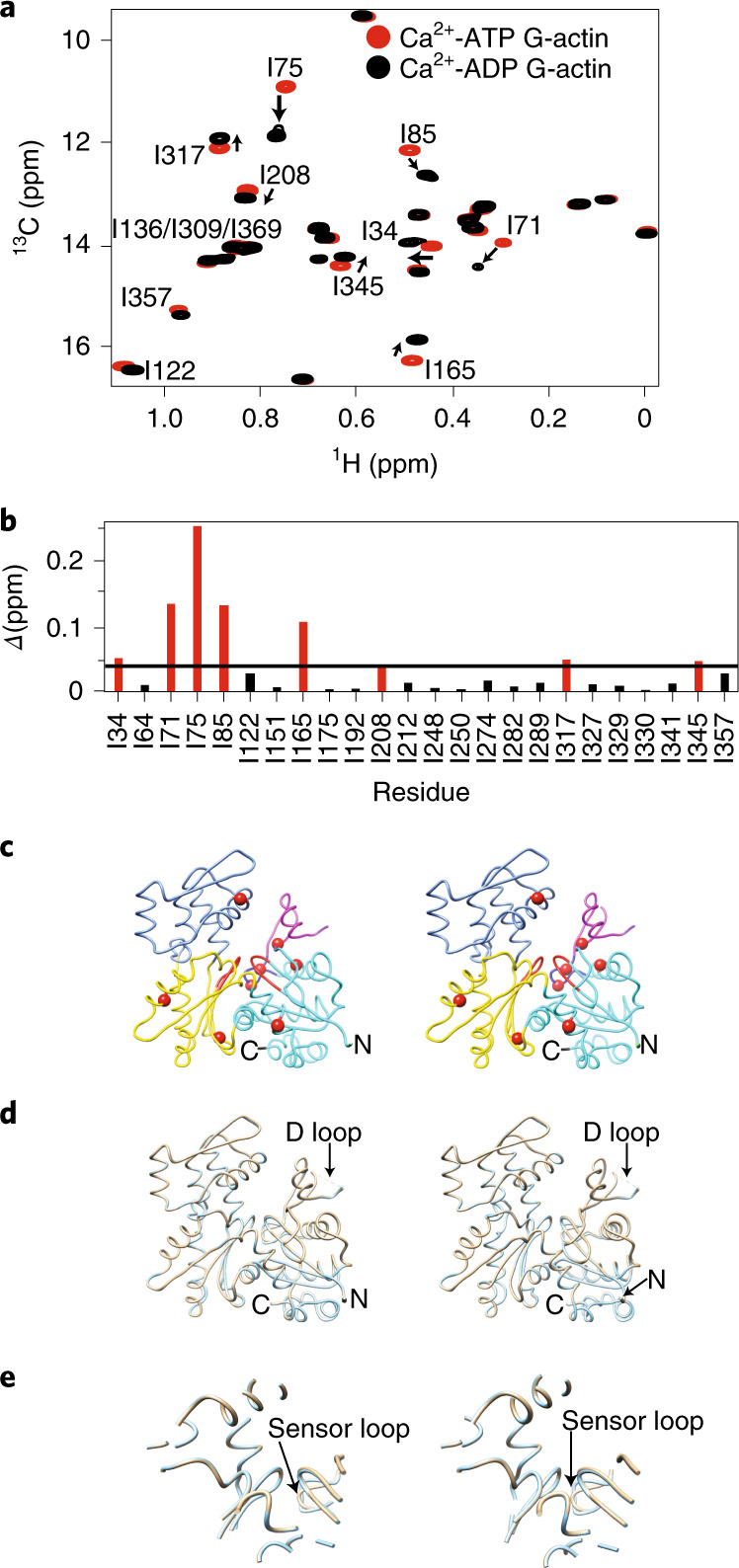
Polymerization of actin into cytoskeletal filaments is coupled to its bound adenine nucleotides. The mechanism by which nucleotide modulates actin functions has not been evident from analyses of ATP- and ADP-bound crystal structures of the actin monomer. We report that NMR chemical shift differences between the two forms are globally distributed. Furthermore, microsecond-millisecond motions are spread throughout the molecule in the ATP form, but largely confined to subdomains 1 and 2, and the nucleotide binding site in the ADP form. Through these motions, the ATP- and ADP-bound forms sample different high-energy conformations. A deafness-causing, fast-nucleating actin mutant populates the high-energy conformer of ATP-actin more than the wild-type protein, suggesting that this conformer may be on the pathway to nucleation. Together, the data suggest a model in which differential sampling of a nucleation-compatible form of the actin monomer may contribute to control of actin filament dynamics by nucleotide.
肌动蛋白聚合形成细胞骨架丝与其结合的腺嘌呤核苷酸有关。核苷酸调节肌动蛋白功能的机制,从分析肌动蛋白单体与 ATP 和 ADP 结合的晶体结构中还不明显。我们报告说,两种形式之间的 NMR 化学位移差异是全局分布的。此外,微秒到毫秒的运动在 ATP 形式中遍布整个分子,但主要局限于结构域 1 和 2 以及 ADP 形式的核苷酸结合位点。通过这些运动,ATP 和 ADP 结合形式可以采样不同的高能构象。一种导致耳聋的快速成核肌动蛋白突变体比野生型蛋白更多地占据 ATP-肌动蛋白的高能构象,这表明该构象可能处于成核途径上。总的来说,这些数据表明,肌动蛋白单体的成核相容形式的差异采样可能有助于核苷酸控制肌动蛋白丝的动力学。