Department of Human Genetics, McGill University, Montreal, Quebec H3A 1B1, Canada.
McGill University Genome Centre, Montreal, Quebec H3A 0G1, Canada.
Genome Res. 2022 May;32(5):825-837. doi: 10.1101/gr.276383.121. Epub 2022 Apr 8.
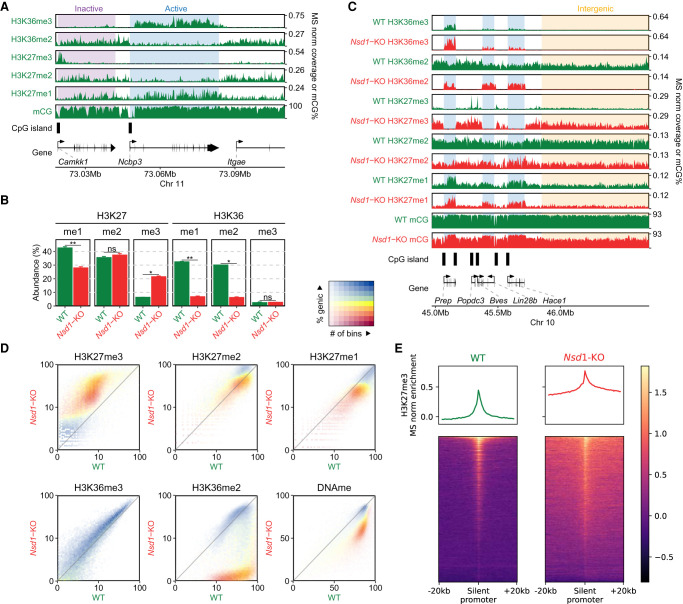
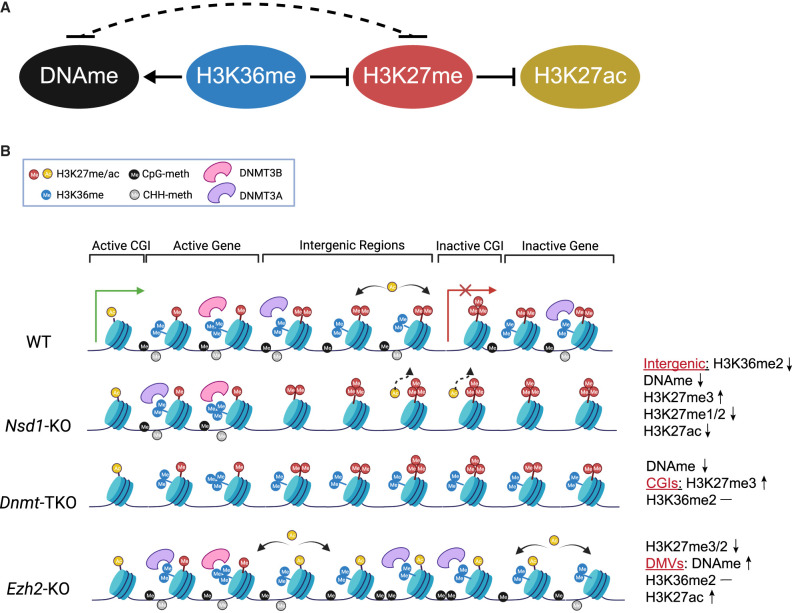
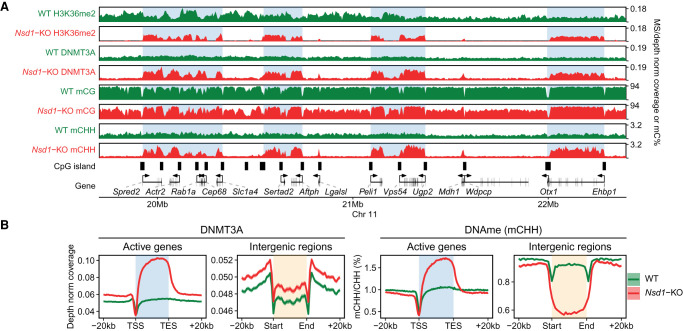
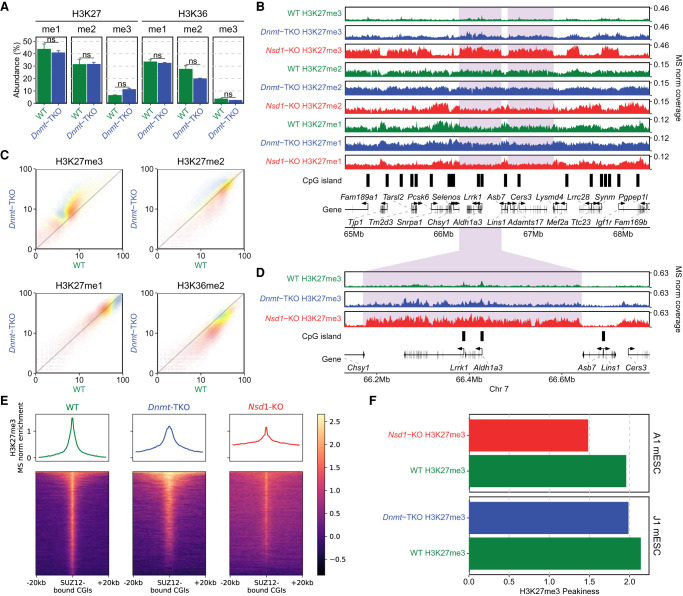
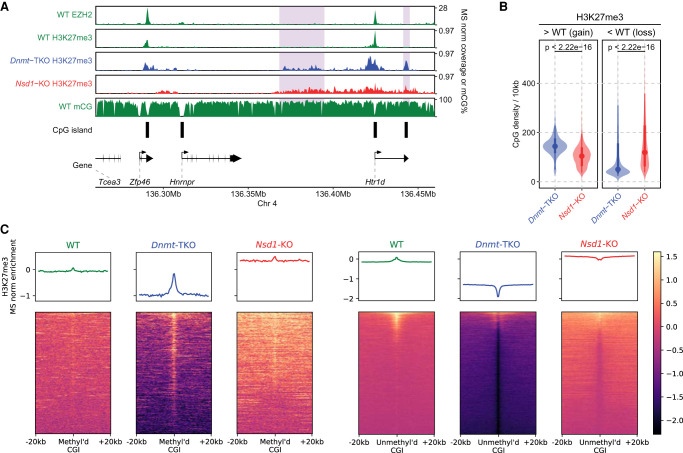
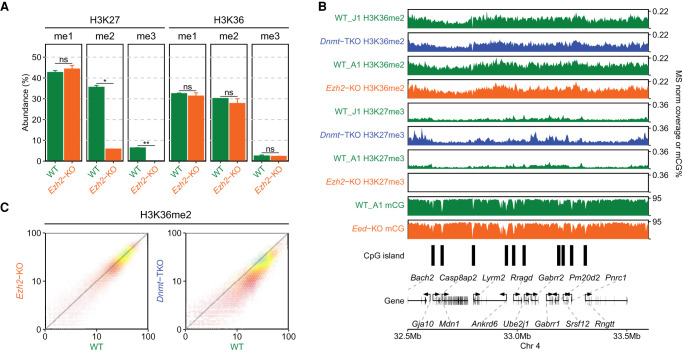
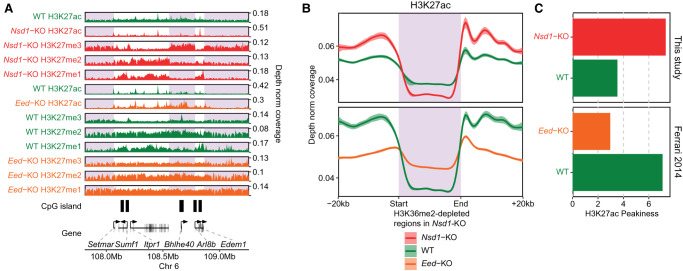
Epigenetic modifications on the chromatin do not occur in isolation. Chromatin-associated proteins and their modification products form a highly interconnected network, and disturbing one component may rearrange the entire system. We see this increasingly clearly in epigenetically dysregulated cancers. It is important to understand the rules governing epigenetic interactions. Here, we use the mouse embryonic stem cell (mESC) model to describe in detail the relationships within the H3K27-H3K36-DNA methylation subnetwork. In particular, we focus on the major epigenetic reorganization caused by deletion of the histone 3 lysine 36 methyltransferase NSD1, which in mESCs deposits nearly all of the intergenic H3K36me2. Although disturbing the H3K27 and DNA methylation (DNAme) components also affects this network to a certain extent, the removal of H3K36me2 has the most drastic effect on the epigenetic landscape, resulting in full intergenic spread of H3K27me3 and a substantial decrease in DNAme. By profiling DNMT3A and CHH methylation (mCHH), we show that H3K36me2 loss upon -KO leads to a massive redistribution of DNMT3A and mCHH away from intergenic regions and toward active gene bodies, suggesting that DNAme reduction is at least in part caused by redistribution of de novo methylation. Additionally, we show that pervasive acetylation of H3K27 is regulated by the interplay of H3K36 and H3K27 methylation. Our analysis highlights the importance of H3K36me2 as a major determinant of the developmental epigenome and provides a framework for further consolidating our knowledge of epigenetic networks.
染色质上的表观遗传修饰并非孤立发生。染色质相关蛋白及其修饰产物形成了一个高度相互关联的网络,干扰一个组件可能会重新排列整个系统。在表观遗传失调的癌症中,我们越来越清楚地看到了这一点。了解控制表观遗传相互作用的规则很重要。在这里,我们使用小鼠胚胎干细胞 (mESC) 模型来详细描述 H3K27-H3K36-DNA 甲基化亚网络内的关系。特别是,我们专注于组蛋白 3 赖氨酸 36 甲基转移酶 NSD1 缺失引起的主要表观遗传重排,在 mESCs 中,该酶几乎沉积了所有基因间的 H3K36me2。尽管干扰 H3K27 和 DNA 甲基化 (DNAme) 成分也会在一定程度上影响这个网络,但 H3K36me2 的去除对表观遗传景观的影响最为剧烈,导致 H3K27me3 在全基因间的广泛扩散和 DNAme 的大量减少。通过对 DNMT3A 和 CHH 甲基化 (mCHH) 的分析,我们发现 -KO 后 H3K36me2 的缺失导致 DNMT3A 和 mCHH 从基因间区域大量重新分配到活跃的基因体,这表明 DNAme 的减少至少部分是由从头甲基化的重新分配引起的。此外,我们还表明,H3K27 的普遍乙酰化受到 H3K36 和 H3K27 甲基化的相互作用调节。我们的分析强调了 H3K36me2 作为发育表观基因组主要决定因素的重要性,并为进一步整合我们对表观遗传网络的知识提供了框架。