Department of Chemistry, University of Massachusetts Amherst, Amherst, United States.
Mass Spectrometry Core Facility, Institute for Applied Life Sciences (IALS), University of Massachusetts Amherst, Amherst, United States.
Elife. 2022 Apr 22;11:e76100. doi: 10.7554/eLife.76100.
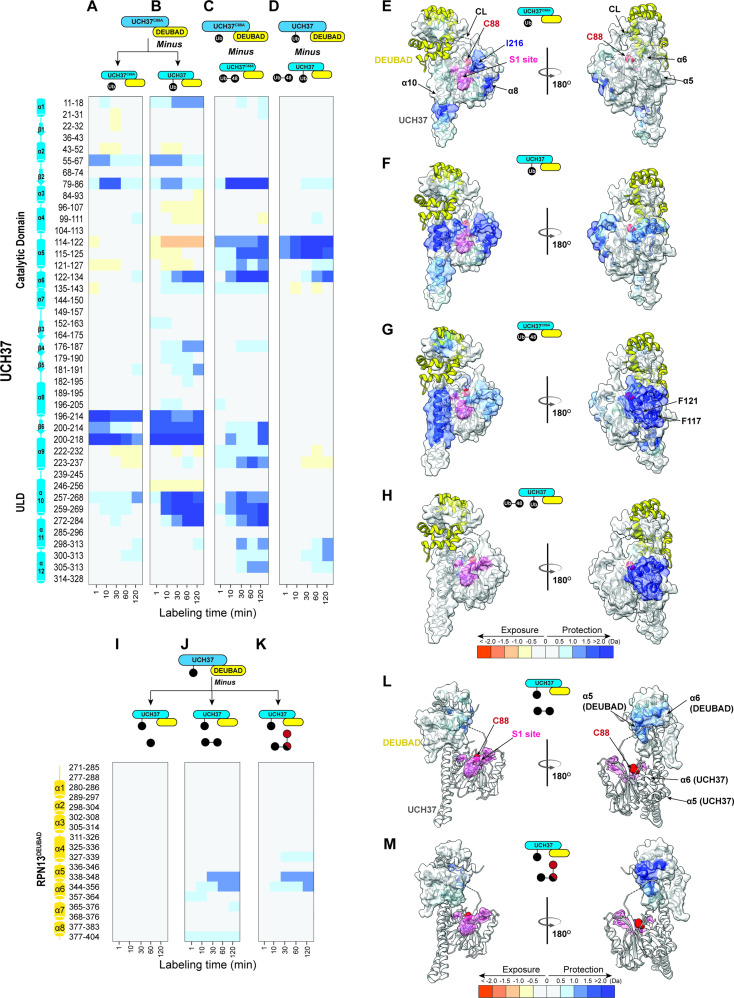
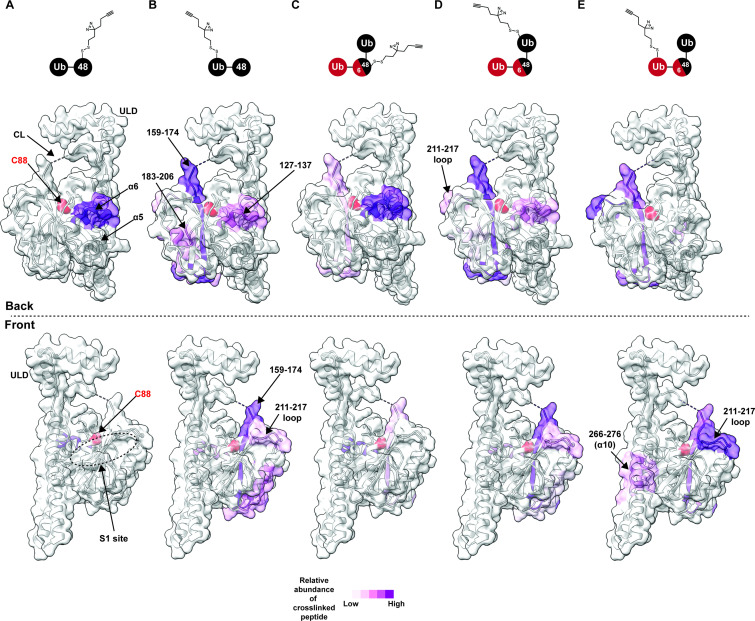
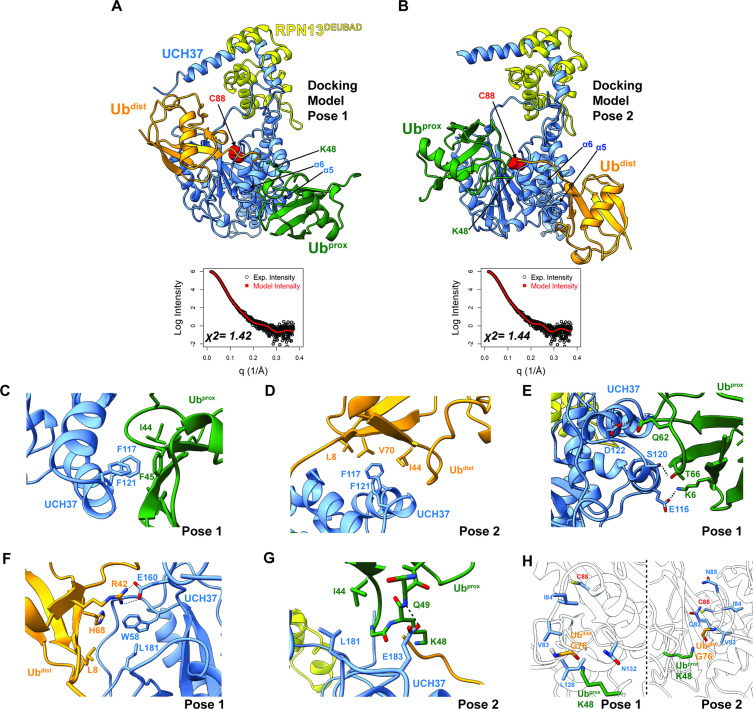
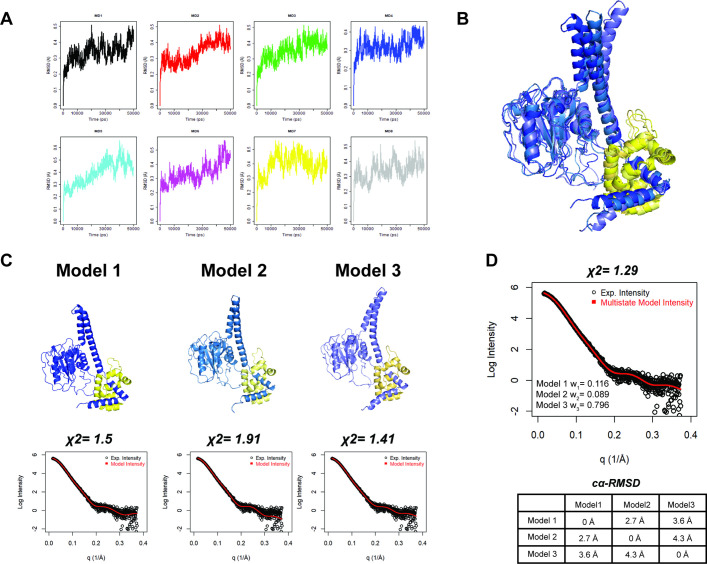
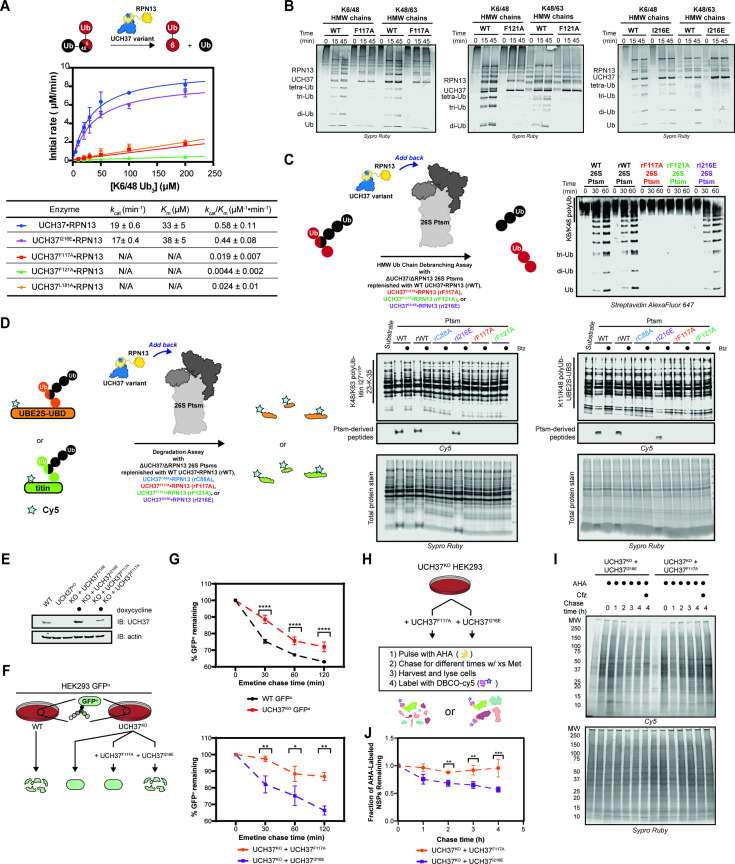
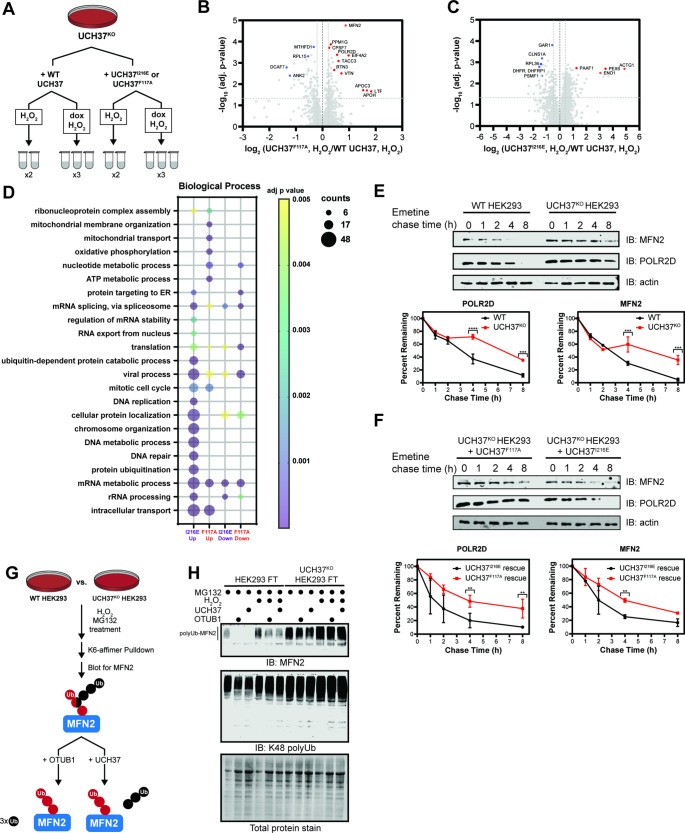
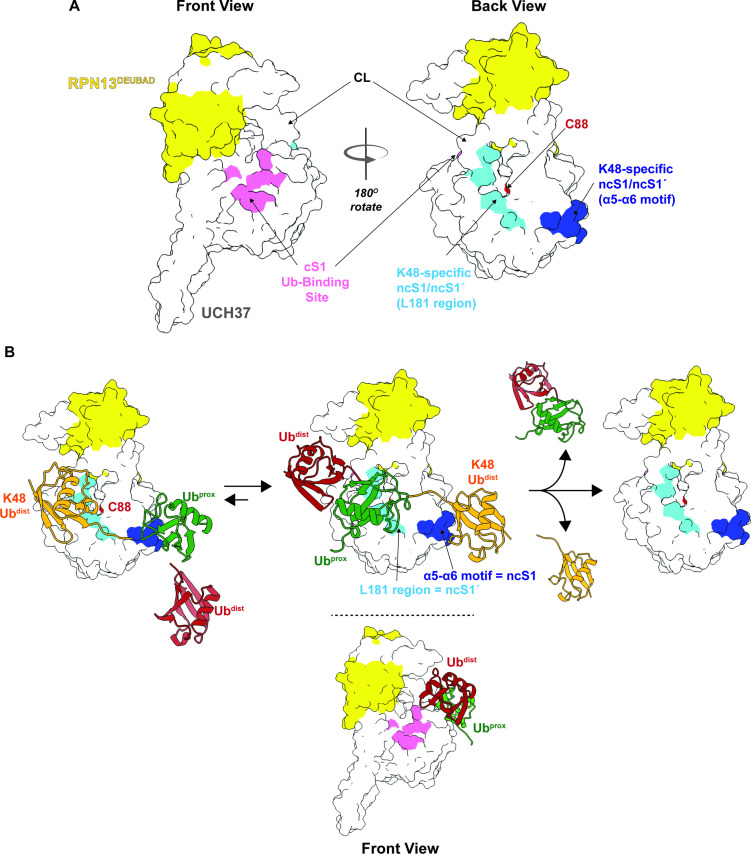
Degradation by the 26 S proteasome is an intricately regulated process fine tuned by the precise nature of ubiquitin modifications attached to a protein substrate. By debranching ubiquitin chains composed of K48 linkages, the proteasome-associated ubiquitin C-terminal hydrolase UCHL5/UCH37 serves as a positive regulator of protein degradation. How UCH37 achieves specificity for K48 chains is unclear. Here, we use a combination of hydrogen-deuterium mass spectrometry, chemical crosslinking, small-angle X-ray scattering, nuclear magnetic resonance (NMR), molecular docking, and targeted mutagenesis to uncover a cryptic K48 ubiquitin (Ub) chain-specific binding site on the opposite face of UCH37 relative to the canonical S1 (cS1) ubiquitin-binding site. Biochemical assays demonstrate the K48 chain-specific binding site is required for chain debranching and proteasome-mediated degradation of proteins modified with branched chains. Using quantitative proteomics, translation shutoff experiments, and linkage-specific affinity tools, we then identify specific proteins whose degradation depends on the debranching activity of UCH37. Our findings suggest that UCH37 and potentially other DUBs could use more than one S1 site to perform different biochemical functions.
26S 蛋白酶体的降解是一个精细调控的过程,受到连接到蛋白质底物上的泛素修饰的精确性质的精细调节。通过去分支由 K48 连接组成的泛素链,与蛋白酶体相关的泛素 C 末端水解酶 UCHL5/UCH37 作为蛋白质降解的正调节剂。UCH37 如何实现对 K48 链的特异性尚不清楚。在这里,我们使用氢氘质谱、化学交联、小角 X 射线散射、核磁共振(NMR)、分子对接和靶向突变来揭示 UCH37 相对于典型 S1(cS1)泛素结合位点在相对的另一侧存在一个隐蔽的 K48 泛素(Ub)链特异性结合位点。生化测定表明,链去分支和带有分支链的蛋白质的蛋白酶体介导的降解需要 K48 链特异性结合位点。使用定量蛋白质组学、翻译关闭实验和连接特异性亲和工具,我们然后鉴定出依赖 UCH37 的去分支活性进行降解的特定蛋白质。我们的发现表明,UCH37 以及可能的其他 DUBs 可以使用不止一个 S1 位点来执行不同的生化功能。