RNA Systems Biochemistry Laboratory, RIKEN Cluster for Pioneering Research, Saitama, Japan.
Cellular Memory Laboratory, RIKEN Cluster for Pioneering Research, Saitama, Japan.
Elife. 2022 Jun 8;11:e72780. doi: 10.7554/eLife.72780.
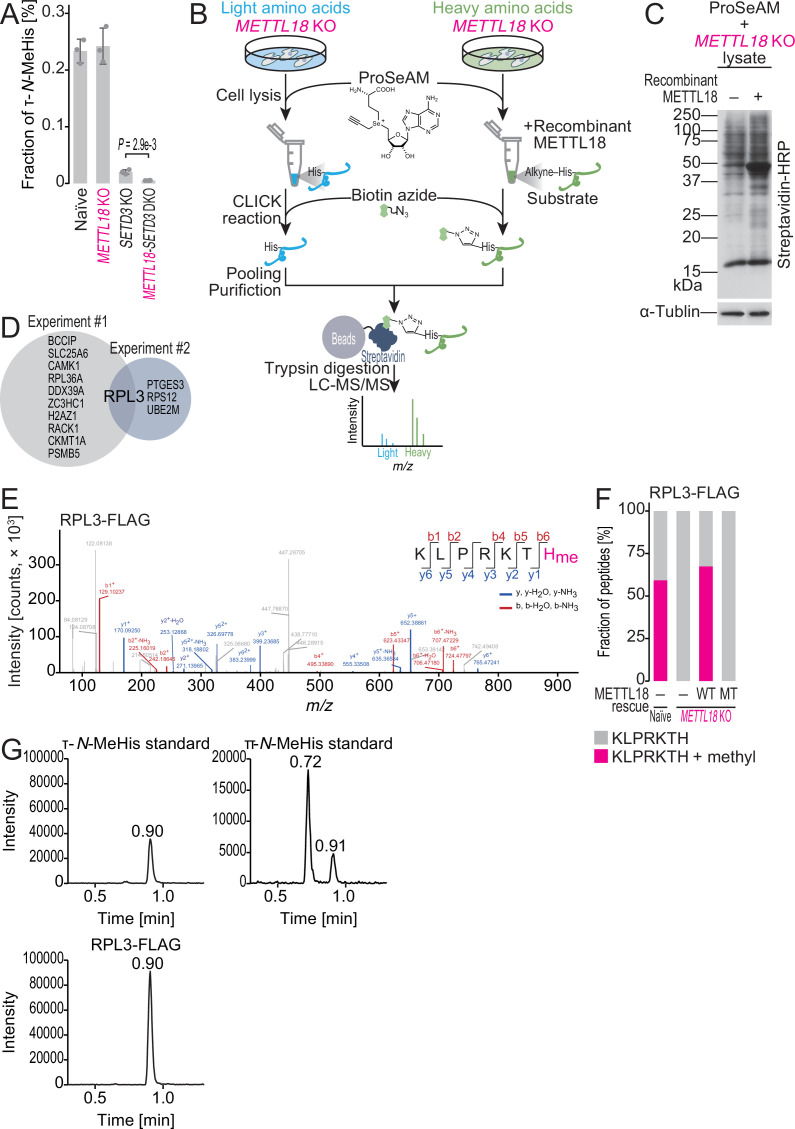
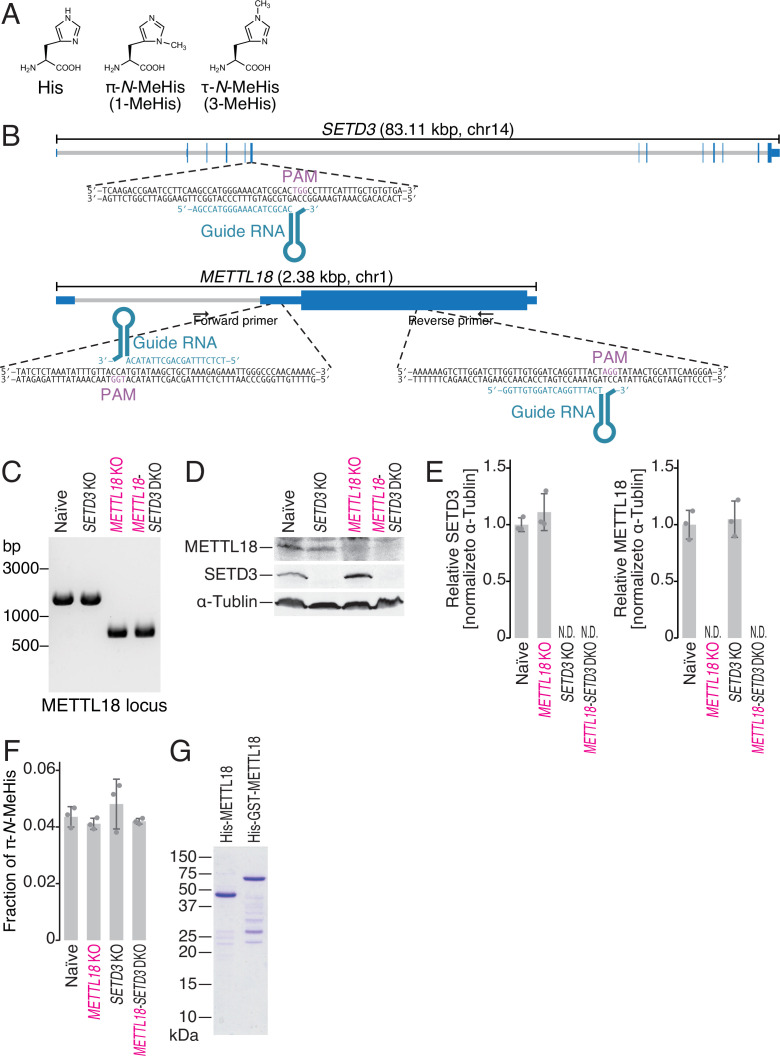
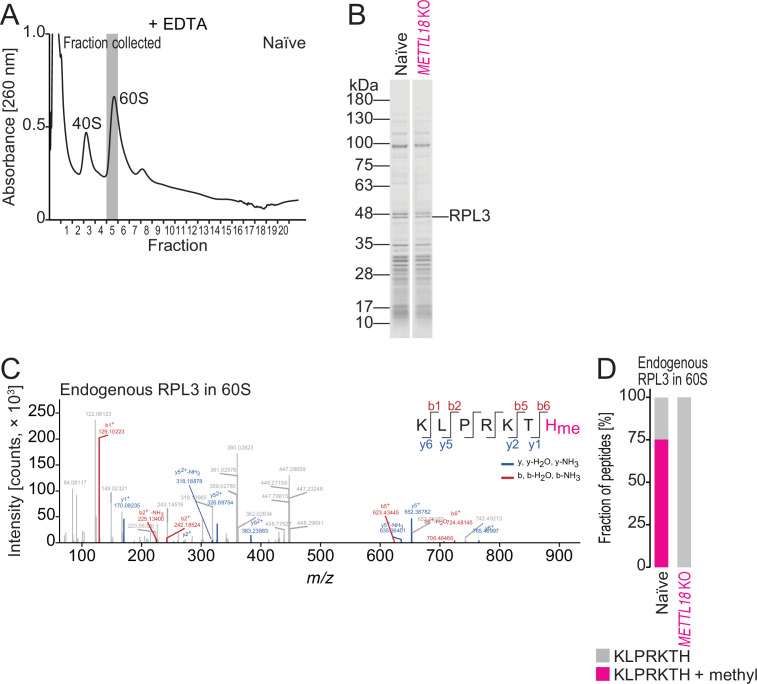
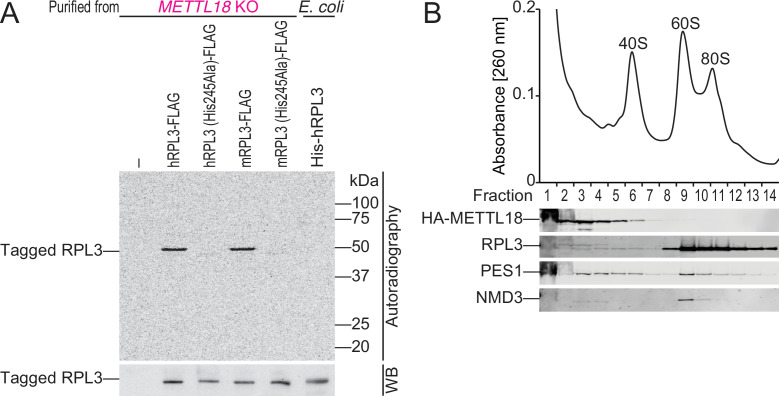
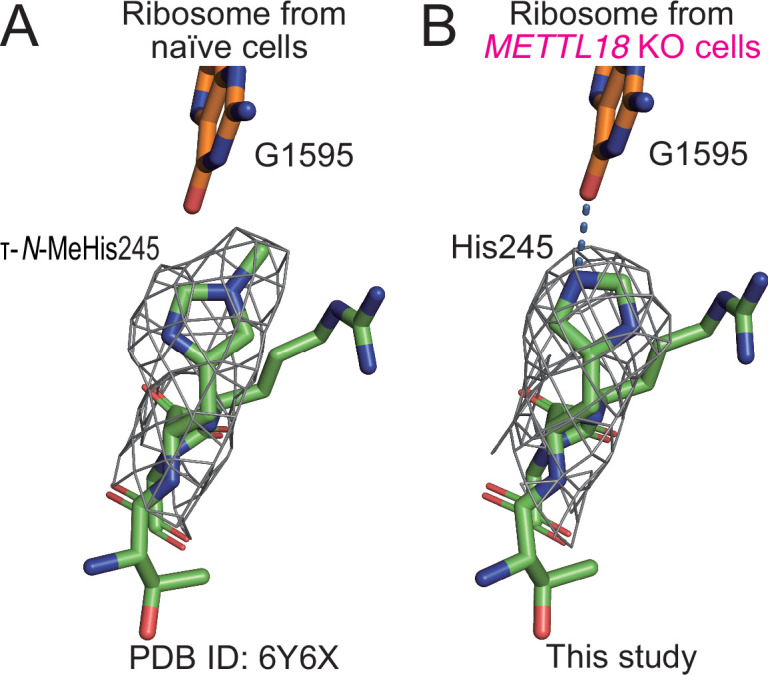
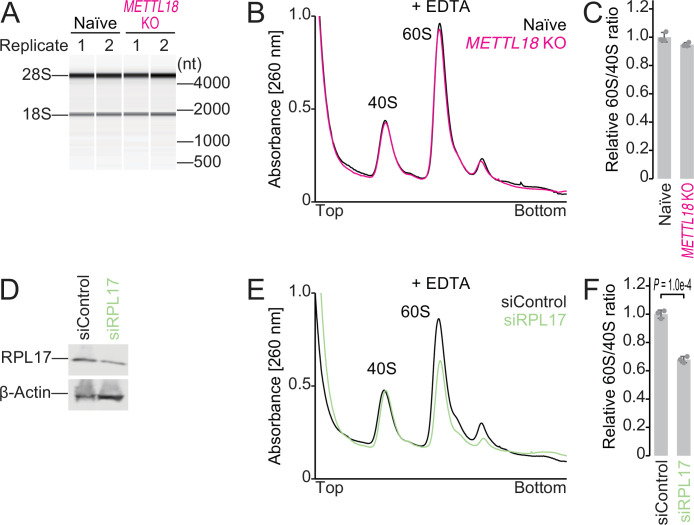
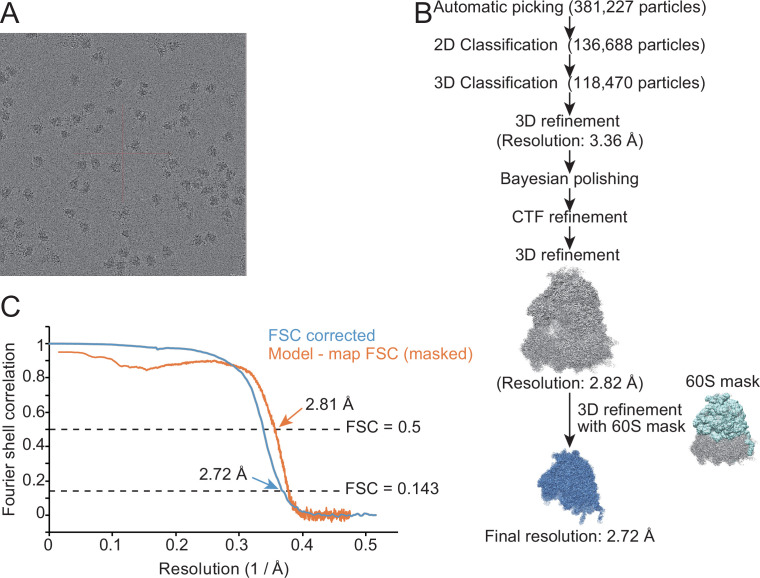
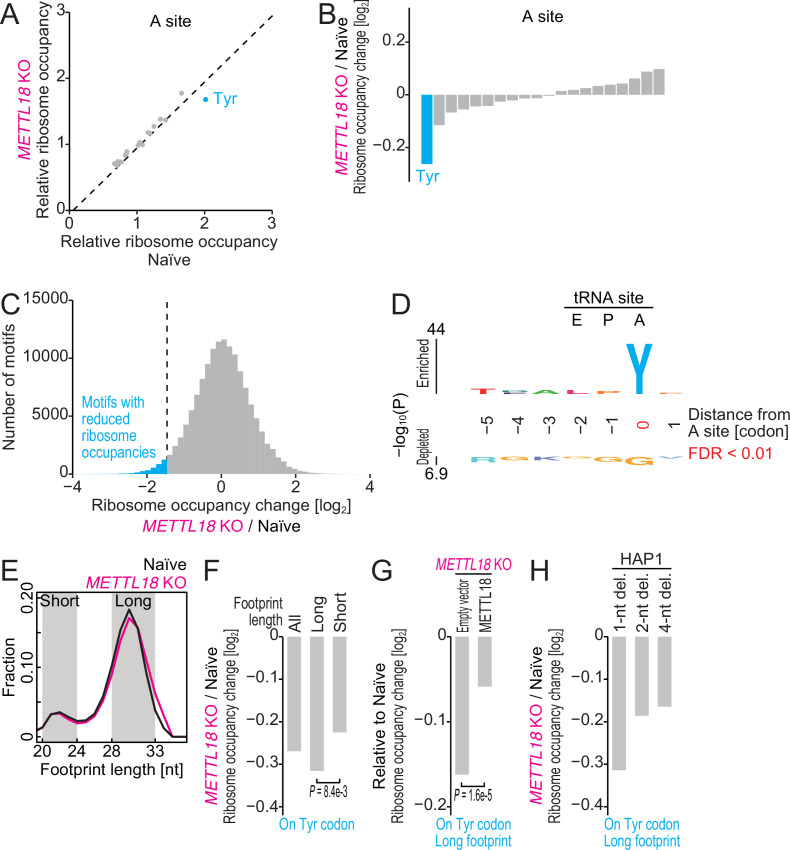
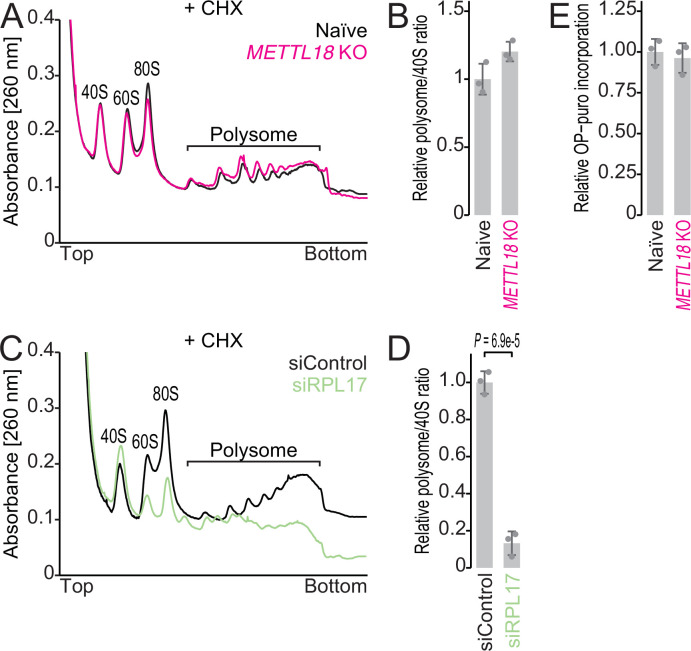
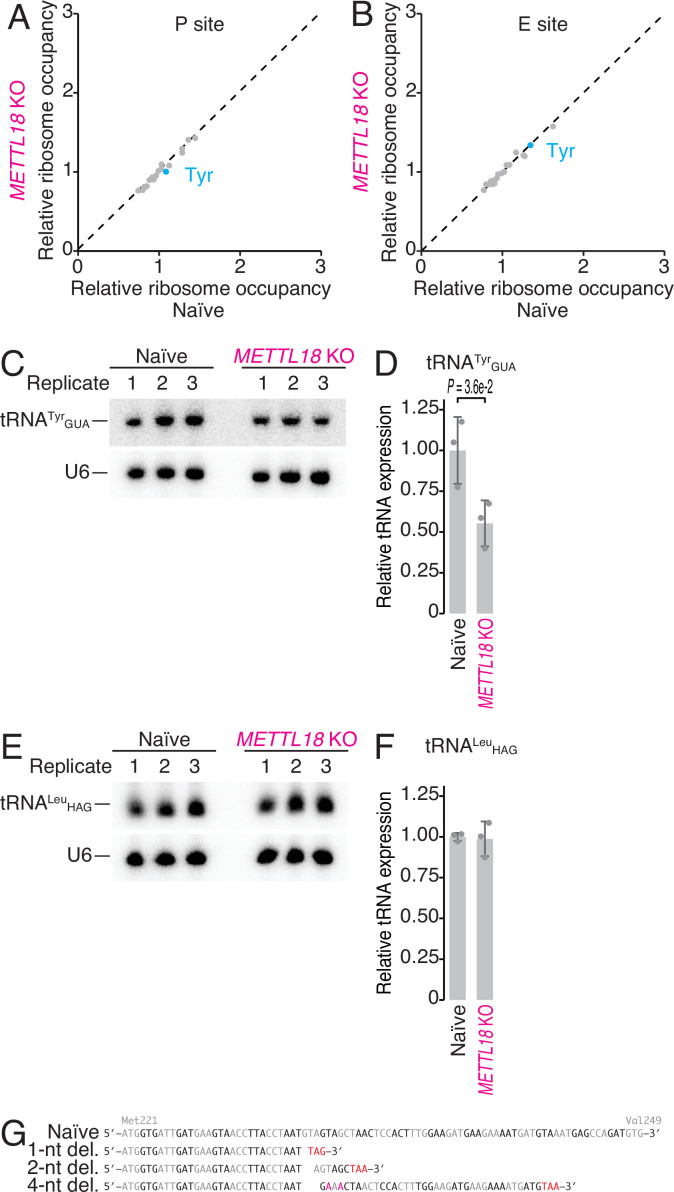
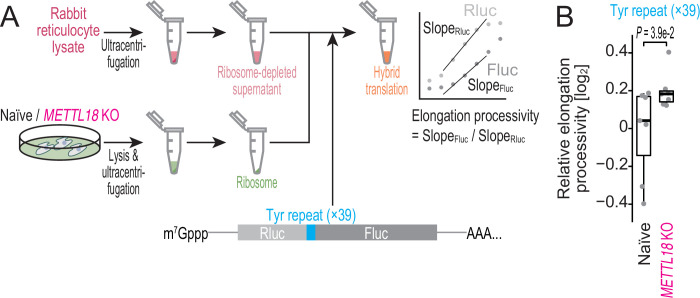
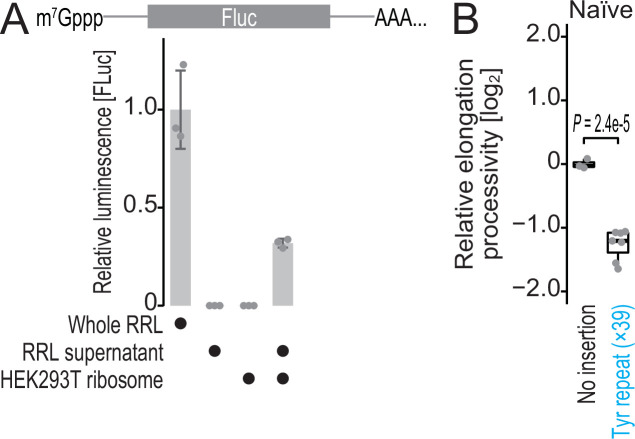
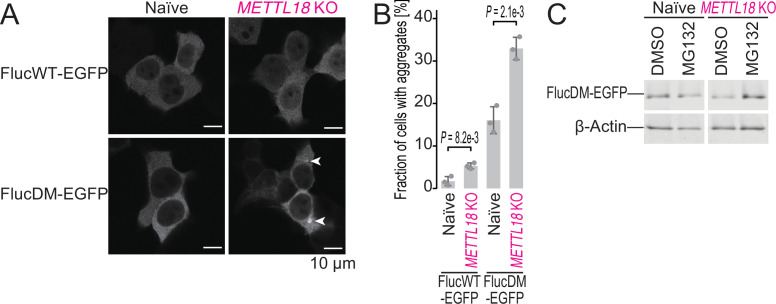
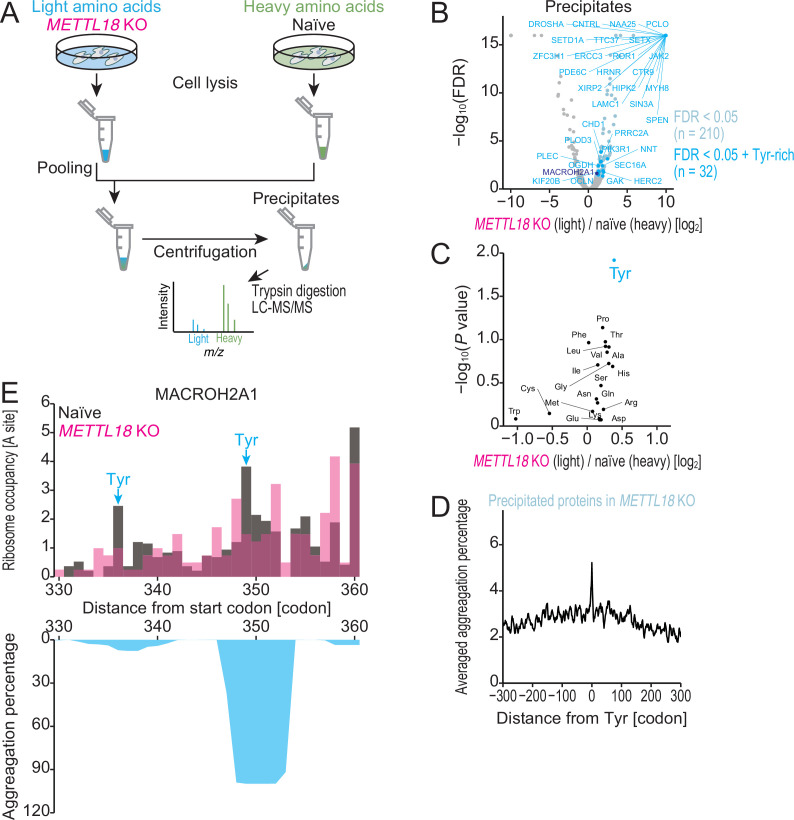
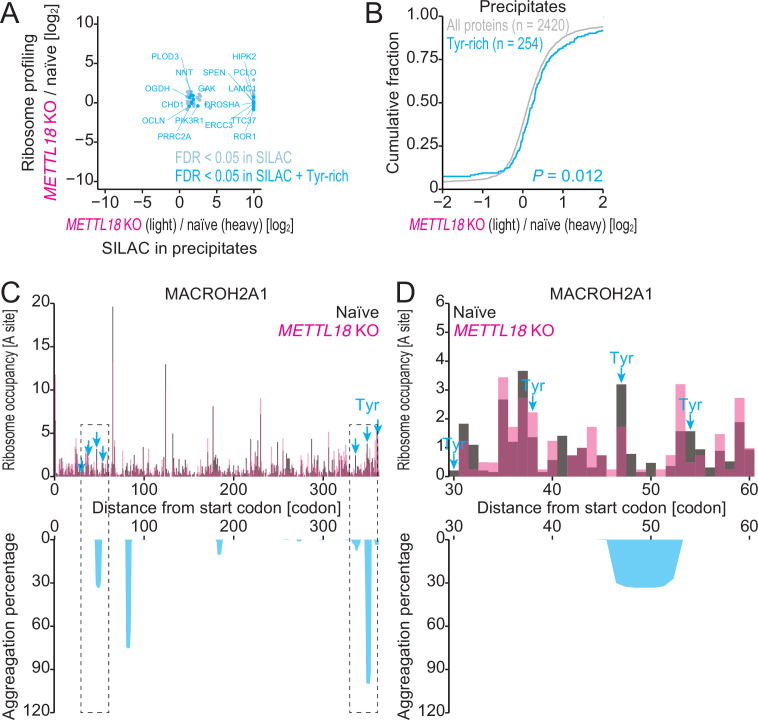
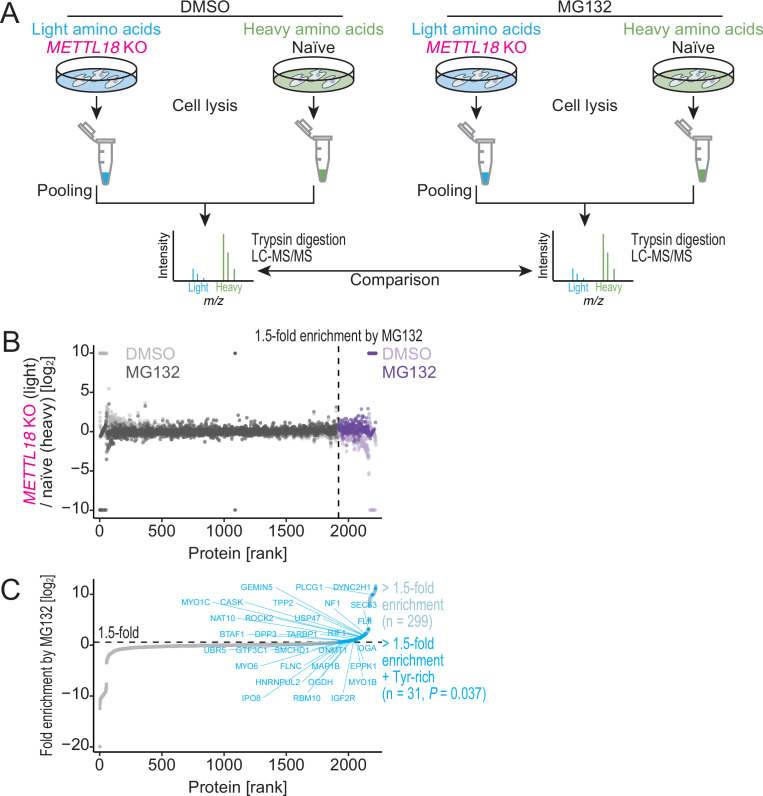
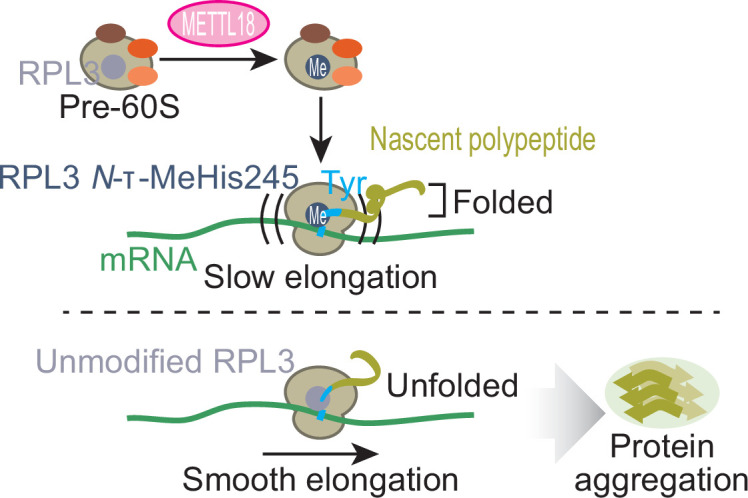
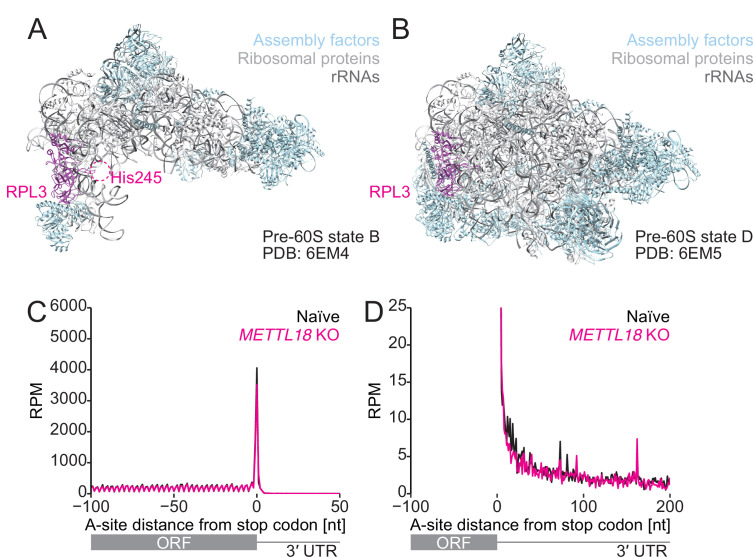
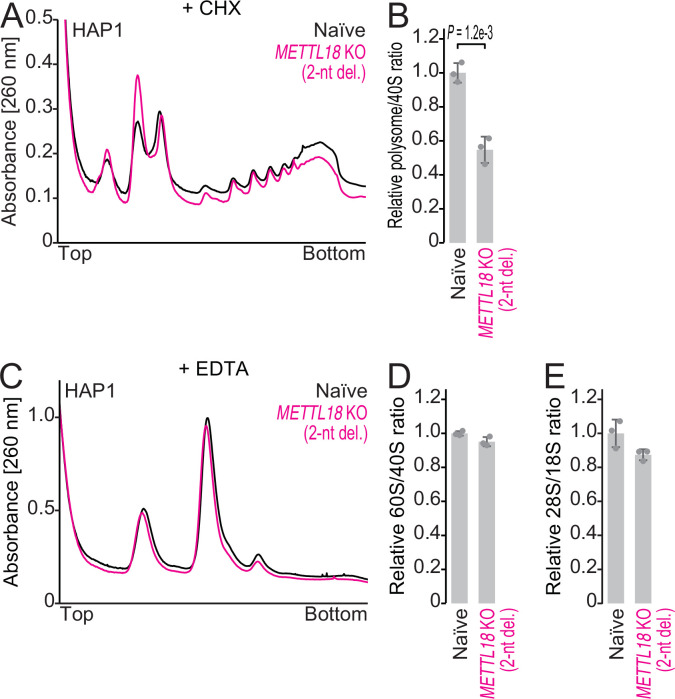
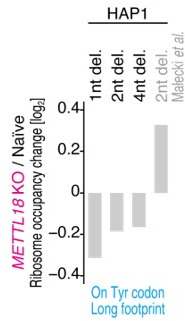
Protein methylation occurs predominantly on lysine and arginine residues, but histidine also serves as a methylation substrate. However, a limited number of enzymes responsible for this modification have been reported. Moreover, the biological role of histidine methylation has remained poorly understood to date. Here, we report that human METTL18 is a histidine methyltransferase for the ribosomal protein RPL3 and that the modification specifically slows ribosome traversal on Tyr codons, allowing the proper folding of synthesized proteins. By performing an in vitro methylation assay with a methyl donor analog and quantitative mass spectrometry, we found that His245 of RPL3 is methylated at the τ- position by METTL18. Structural comparison of the modified and unmodified ribosomes showed stoichiometric modification and suggested a role in translation reactions. Indeed, genome-wide ribosome profiling and an in vitro translation assay revealed that translation elongation at Tyr codons was suppressed by RPL3 methylation. Because the slower elongation provides enough time for nascent protein folding, RPL3 methylation protects cells from the cellular aggregation of Tyr-rich proteins. Our results reveal histidine methylation as an example of a ribosome modification that ensures proteome integrity in cells.
蛋白质甲基化主要发生在赖氨酸和精氨酸残基上,但组氨酸也可作为甲基化的底物。然而,迄今为止,已经报道了负责这种修饰的酶的数量有限。此外,组氨酸甲基化的生物学作用至今仍知之甚少。在这里,我们报告人类 METTL18 是核糖体蛋白 RPL3 的组氨酸甲基转移酶,该修饰特异性地减缓了 Tyr 密码子上的核糖体穿越,从而使合成的蛋白质能够正确折叠。通过使用甲基供体类似物进行体外甲基化测定和定量质谱分析,我们发现 RPL3 的 His245 被 METTL18 在τ-位置甲基化。修饰和未修饰核糖体的结构比较表明存在化学计量修饰,并提示其在翻译反应中起作用。事实上,全基因组核糖体图谱分析和体外翻译测定表明,RPL3 甲基化抑制了 Tyr 密码子处的翻译延伸。由于较慢的延伸为新生蛋白折叠提供了足够的时间,因此 RPL3 甲基化可防止富含 Tyr 的蛋白质在细胞中聚集。我们的研究结果揭示了组氨酸甲基化作为核糖体修饰的一个例子,它可确保细胞中蛋白质组的完整性。