National Institute of Neurological Disorders and Stroke, Bethesda, MD, USA.
Department of Physiology and Pharmacology, Sackler Faculty of Medicine, Tel Aviv University, 69978, Ramat Aviv, Israel.
Nat Commun. 2022 Jun 27;13(1):3697. doi: 10.1038/s41467-022-31286-4.
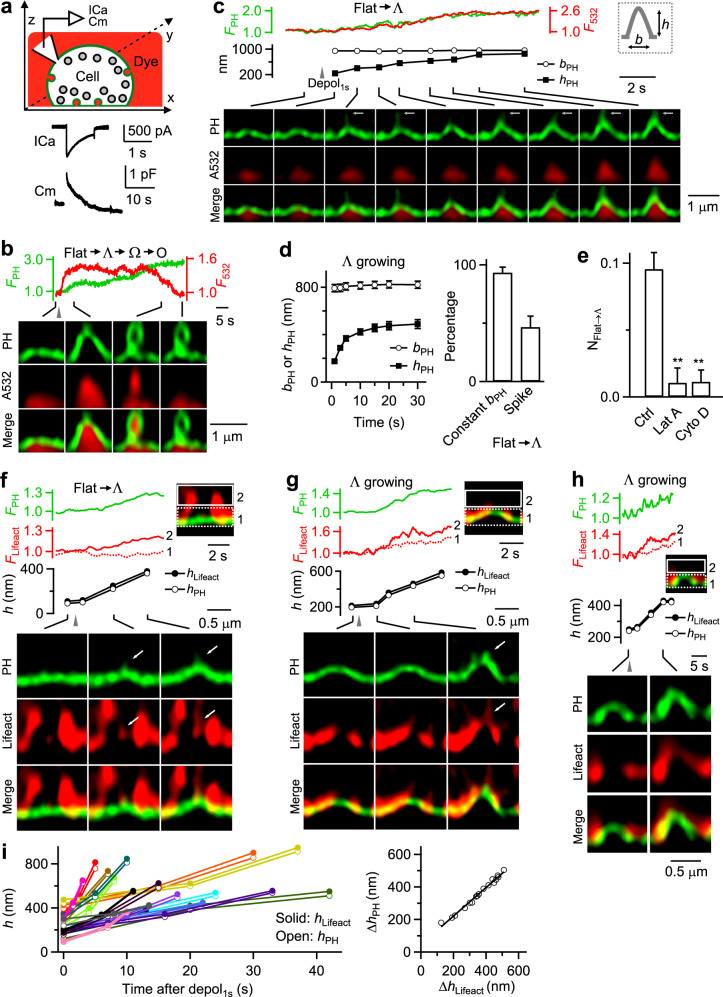
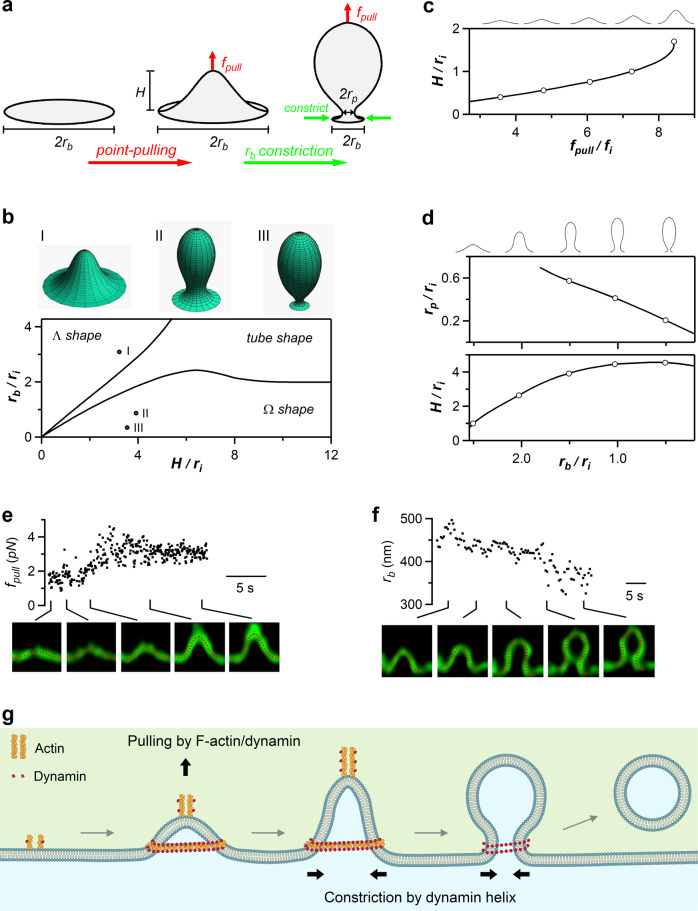
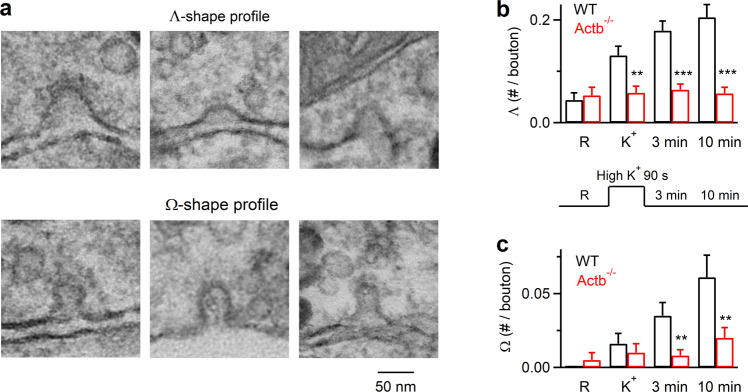
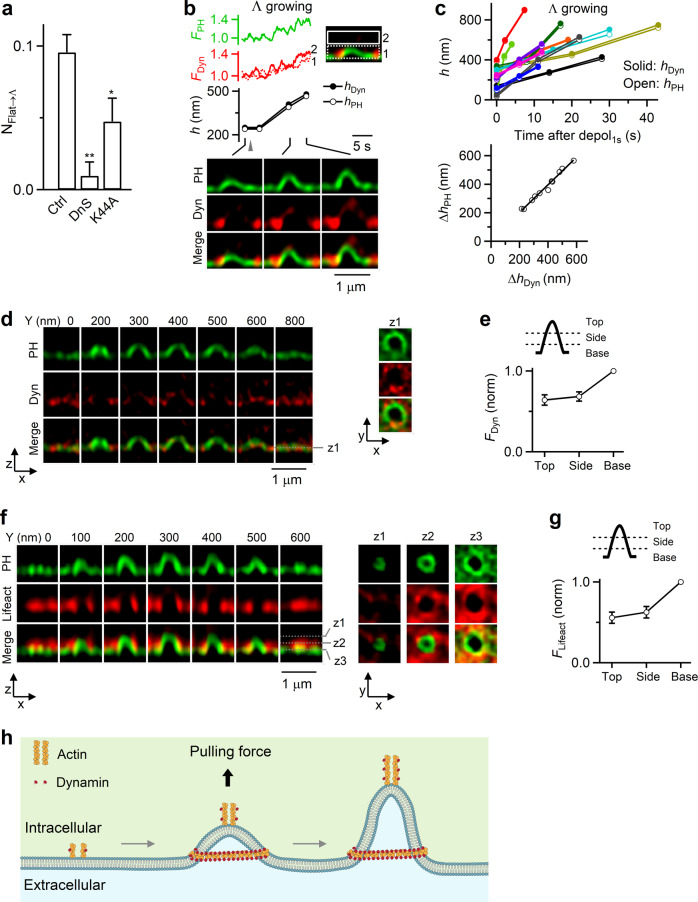
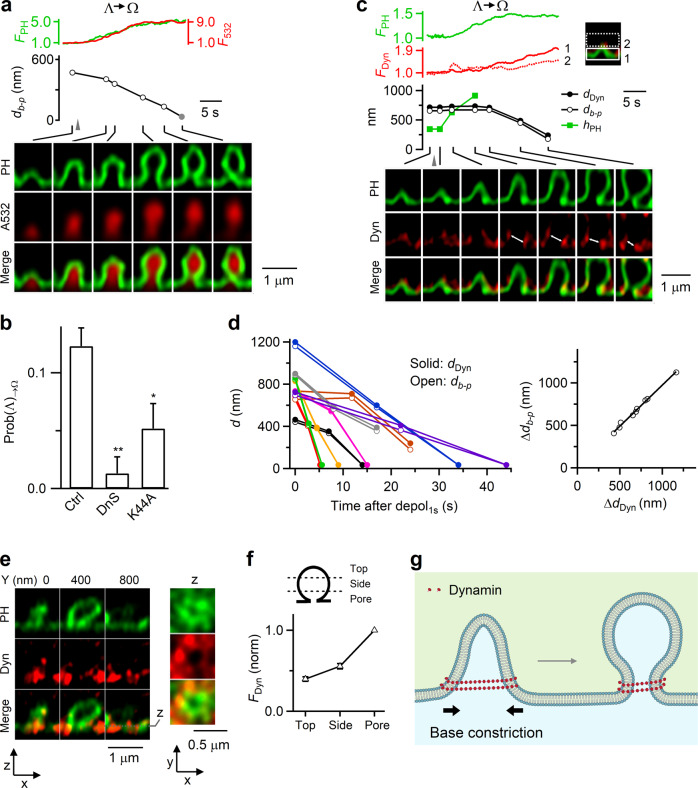
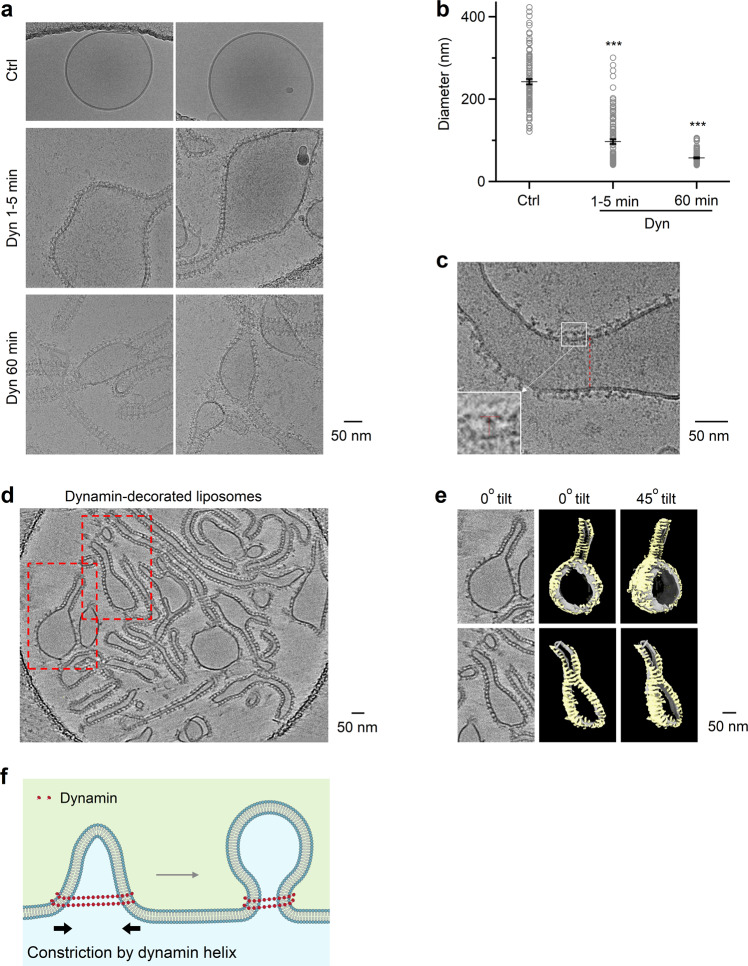
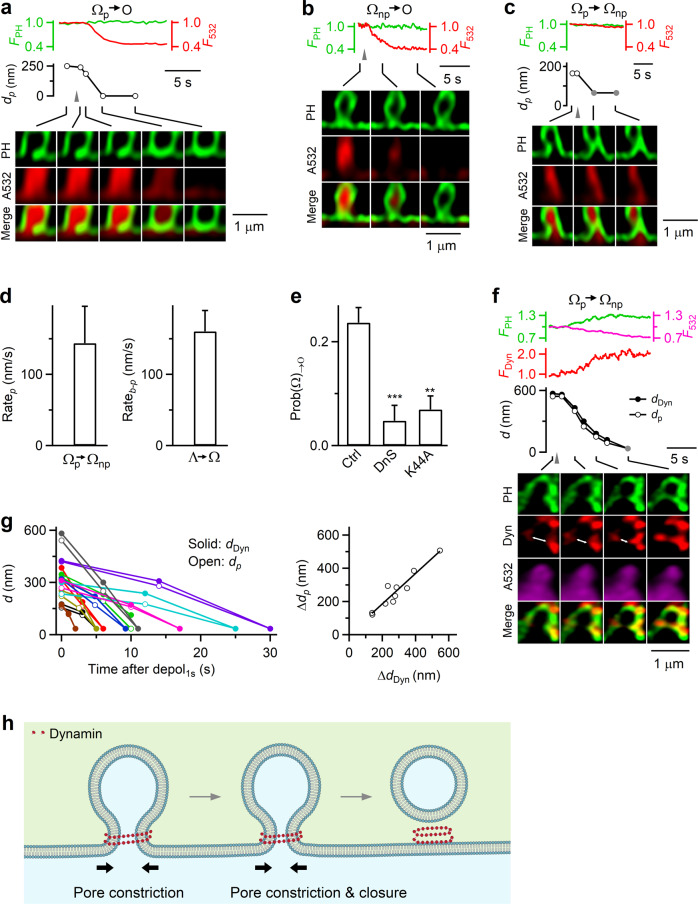
Membrane budding entails forces to transform flat membrane into vesicles essential for cell survival. Accumulated studies have identified coat-proteins (e.g., clathrin) as potential budding factors. However, forces mediating many non-coated membrane buddings remain unclear. By visualizing proteins in mediating endocytic budding in live neuroendocrine cells, performing in vitro protein reconstitution and physical modeling, we discovered how non-coated-membrane budding is mediated: actin filaments and dynamin generate a pulling force transforming flat membrane into Λ-shape; subsequently, dynamin helices surround and constrict Λ-profile's base, transforming Λ- to Ω-profile, and then constrict Ω-profile's pore, converting Ω-profiles to vesicles. These mechanisms control budding speed, vesicle size and number, generating diverse endocytic modes differing in these parameters. Their impact is widespread beyond secretory cells, as the unexpectedly powerful functions of dynamin and actin, previously thought to mediate fission and overcome tension, respectively, may contribute to many dynamin/actin-dependent non-coated-membrane buddings, coated-membrane buddings, and other membrane remodeling processes.
膜泡出芽需要将平面膜转化为泡囊的力,这对于细胞存活至关重要。大量研究已经确定了衣被蛋白(如网格蛋白)是潜在的出芽因子。然而,介导许多非包被膜出芽的力仍不清楚。通过在活体神经内分泌细胞中可视化介导内吞作用出芽的蛋白质,进行体外蛋白质重组和物理建模,我们发现了非包被膜出芽是如何被介导的:肌动蛋白丝和动力蛋白产生拉力,将平面膜转化为 Λ 形;随后,动力蛋白螺旋围绕并收缩 Λ 形的底部,将 Λ 形转化为 Ω 形,然后收缩 Ω 形的孔,将 Ω 形转化为泡囊。这些机制控制着出芽的速度、囊泡的大小和数量,产生了在这些参数上不同的多种内吞模式。这些机制的影响不仅限于分泌细胞,因为动力蛋白和肌动蛋白的功能出乎意料地强大,以前被认为分别介导分裂和克服张力,它们可能有助于许多依赖动力蛋白/肌动蛋白的非包被膜出芽、包被膜出芽和其他膜重塑过程。