MRC Laboratory of Molecular Biology, Francis Crick Avenue, Cambridge CB2 0QH, UK.
Division of Virology, Department of Pathology, University of Cambridge, Addenbrooke's Hospital, Cambridge CB2 0QQ, UK.
Viruses. 2022 Jul 23;14(8):1610. doi: 10.3390/v14081610.
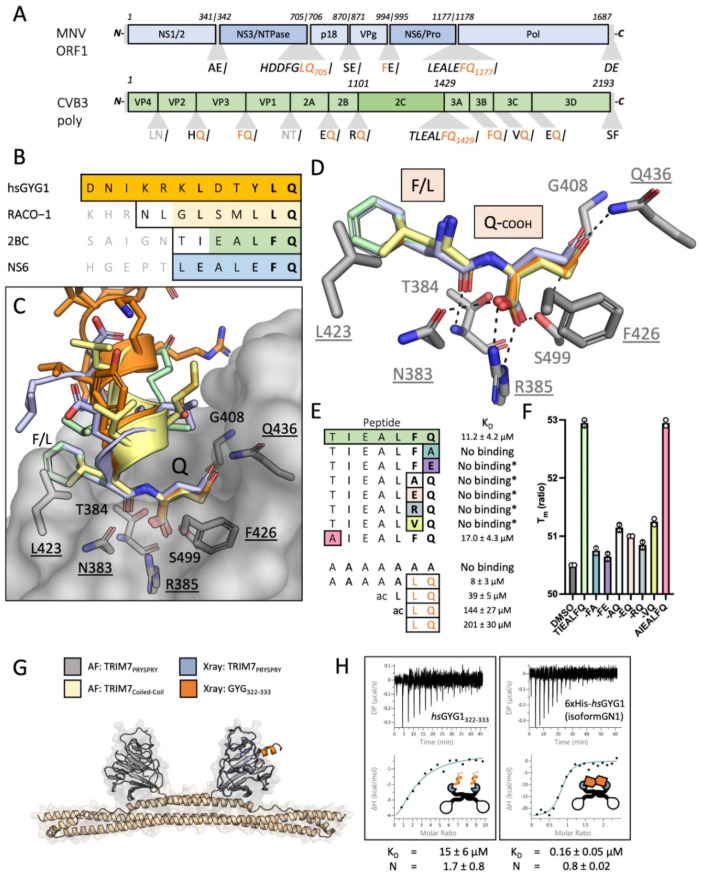
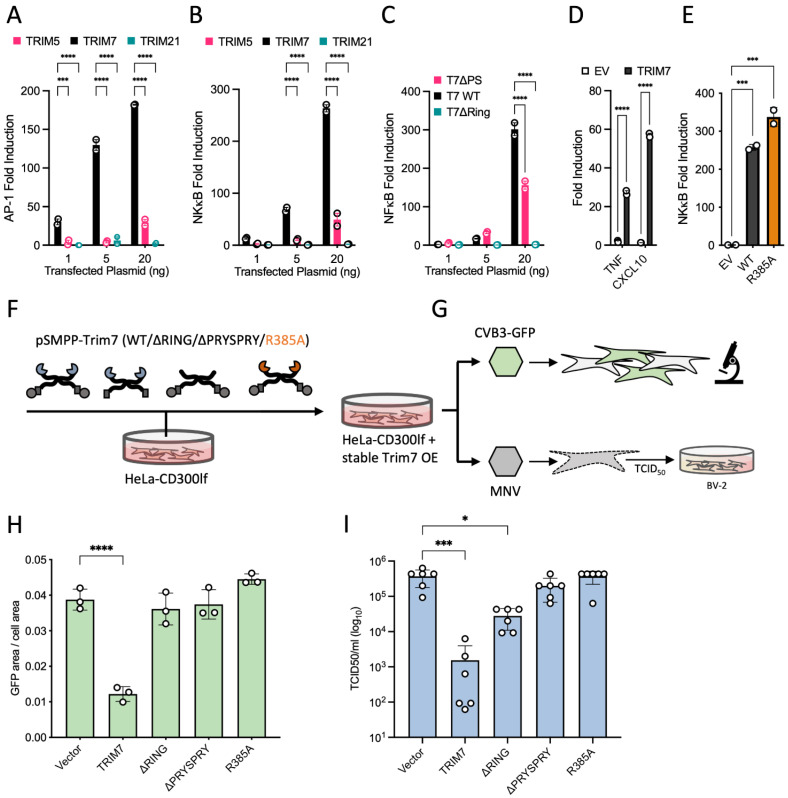
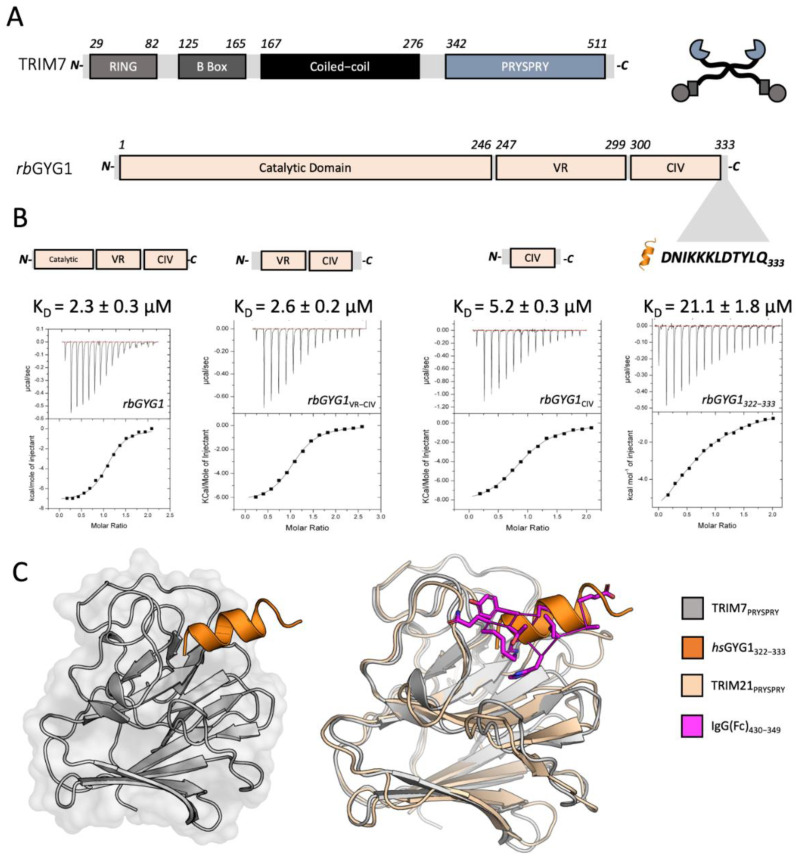
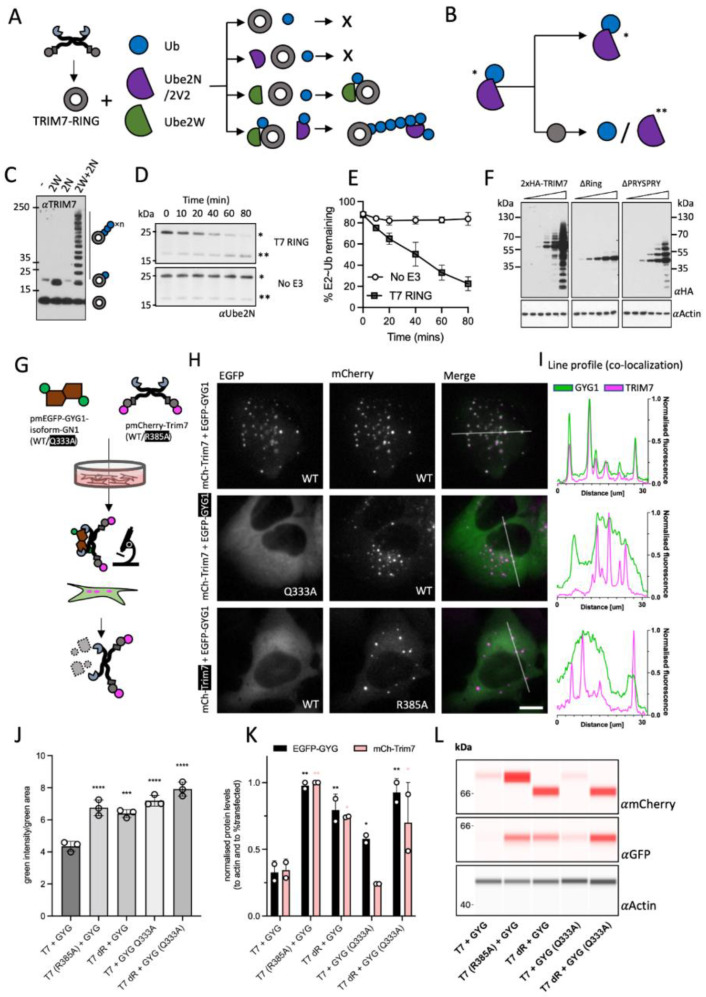
TRIM7 catalyzes the ubiquitination of multiple substrates with unrelated biological functions. This cross-reactivity is at odds with the specificity usually displayed by enzymes, including ubiquitin ligases. Here we show that TRIM7's extreme substrate promiscuity is due to a highly unusual binding mechanism, in which the PRYSPRY domain captures any ligand with a C-terminal helix that terminates in a hydrophobic residue followed by a glutamine. Many of the non-structural proteins found in RNA viruses contain C-terminal glutamines as a result of polyprotein cleavage by 3C protease. This viral processing strategy generates novel substrates for TRIM7 and explains its ability to inhibit Coxsackie virus and norovirus replication. In addition to viral proteins, cellular proteins such as glycogenin have evolved C-termini that make them a TRIM7 substrate. The 'helix-ΦQ' degron motif recognized by TRIM7 is reminiscent of the N-end degron system and is found in ~1% of cellular proteins. These features, together with TRIM7's restricted tissue expression and lack of immune regulation, suggest that viral restriction may not be its physiological function.
TRIM7 催化具有不同生物学功能的多种底物的泛素化。这种交叉反应与通常由酶(包括泛素连接酶)显示的特异性不一致。在这里,我们表明 TRIM7 的极端底物混杂性是由于一种非常不寻常的结合机制,其中 PRYSPRY 结构域捕获任何具有 C 末端螺旋的配体,该螺旋在疏水残基后终止,然后是谷氨酰胺。许多在 RNA 病毒中发现的非结构蛋白含有 C 末端谷氨酰胺,这是 3C 蛋白酶对多蛋白切割的结果。这种病毒加工策略为 TRIM7 生成了新的底物,并解释了其抑制柯萨奇病毒和诺如病毒复制的能力。除了病毒蛋白外,细胞蛋白(如糖原合酶)的 C 末端也发生了进化,使它们成为 TRIM7 的底物。TRIM7 识别的“螺旋-ΦQ”降解基序让人联想到 N 端降解基序,并且在大约 1%的细胞蛋白中发现。这些特征,加上 TRIM7 受限的组织表达和缺乏免疫调节,表明病毒限制可能不是其生理功能。