Centro Nacional de Investigaciones Cardiovasculares, Madrid, Spain.
CIBER de Enfermedades Cardiovasculares, Madrid, Spain.
Elife. 2022 Aug 16;11:e75250. doi: 10.7554/eLife.75250.
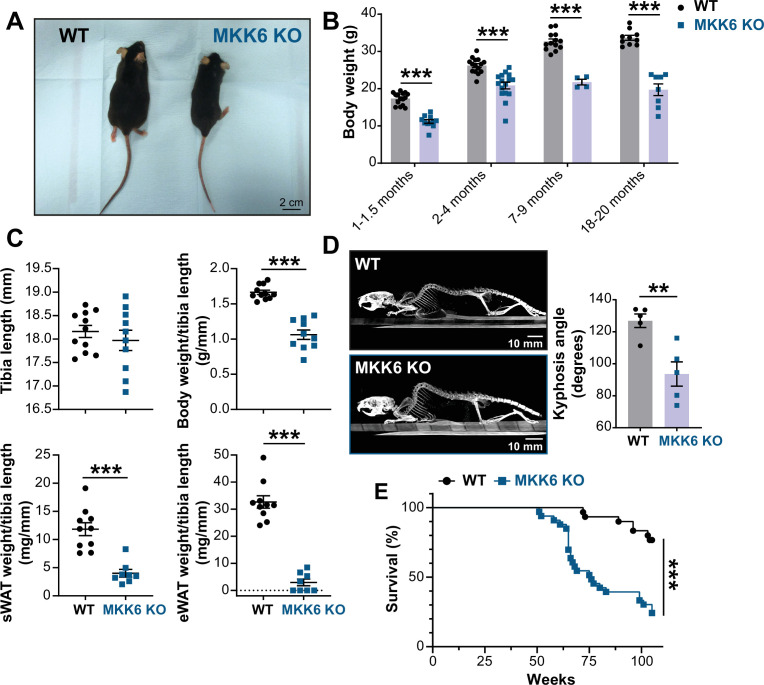
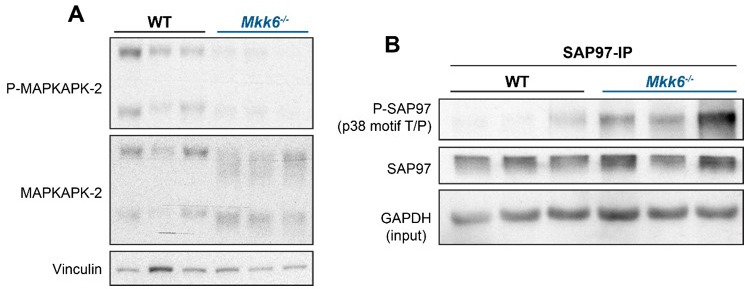
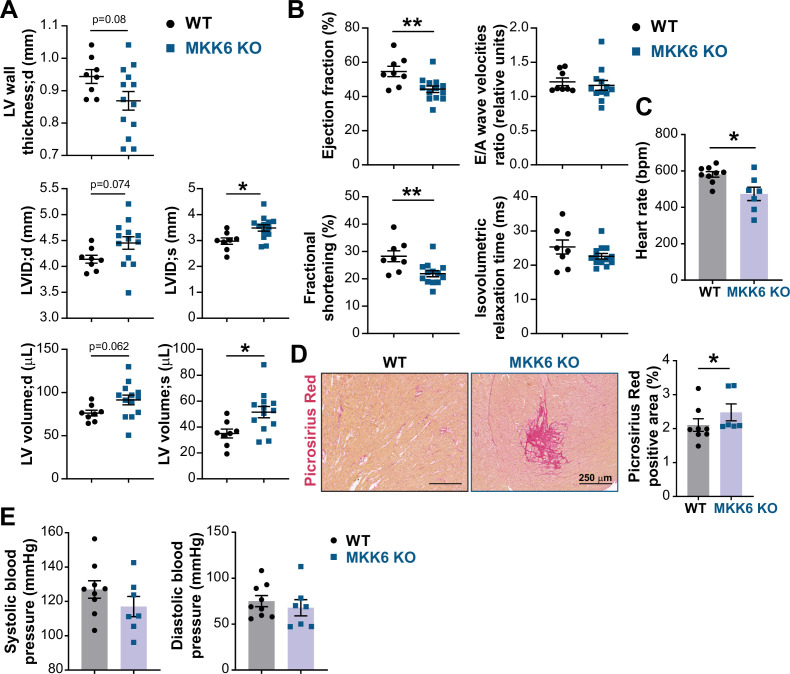
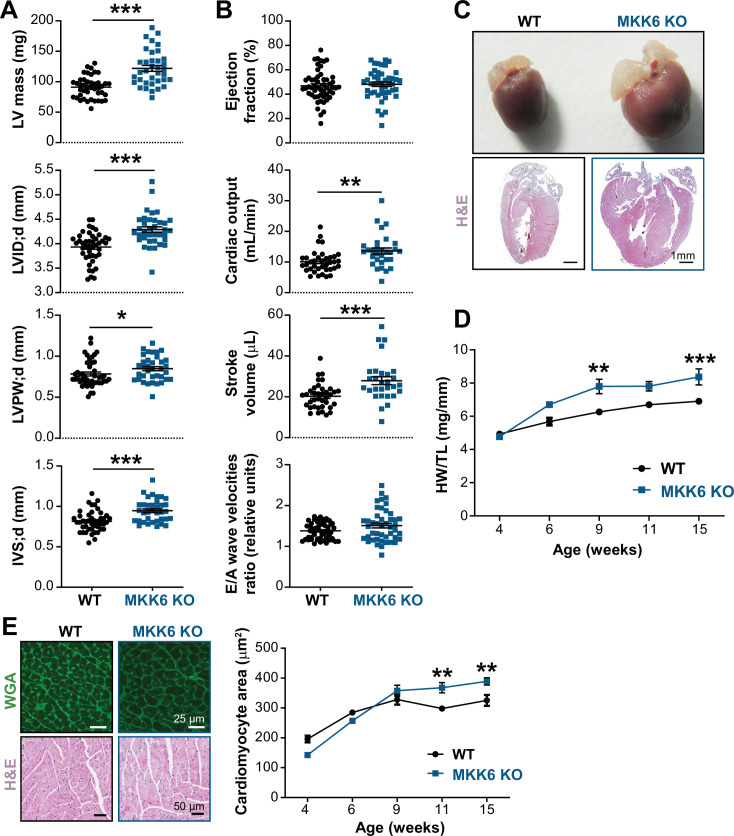
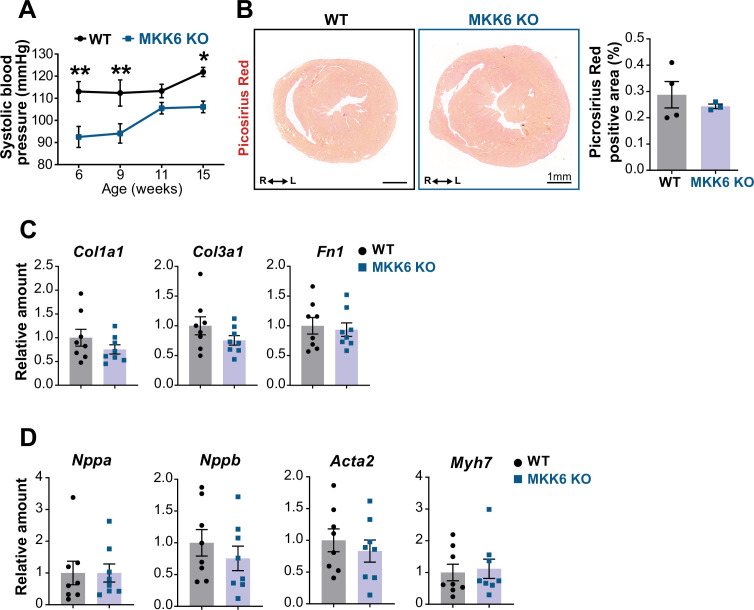
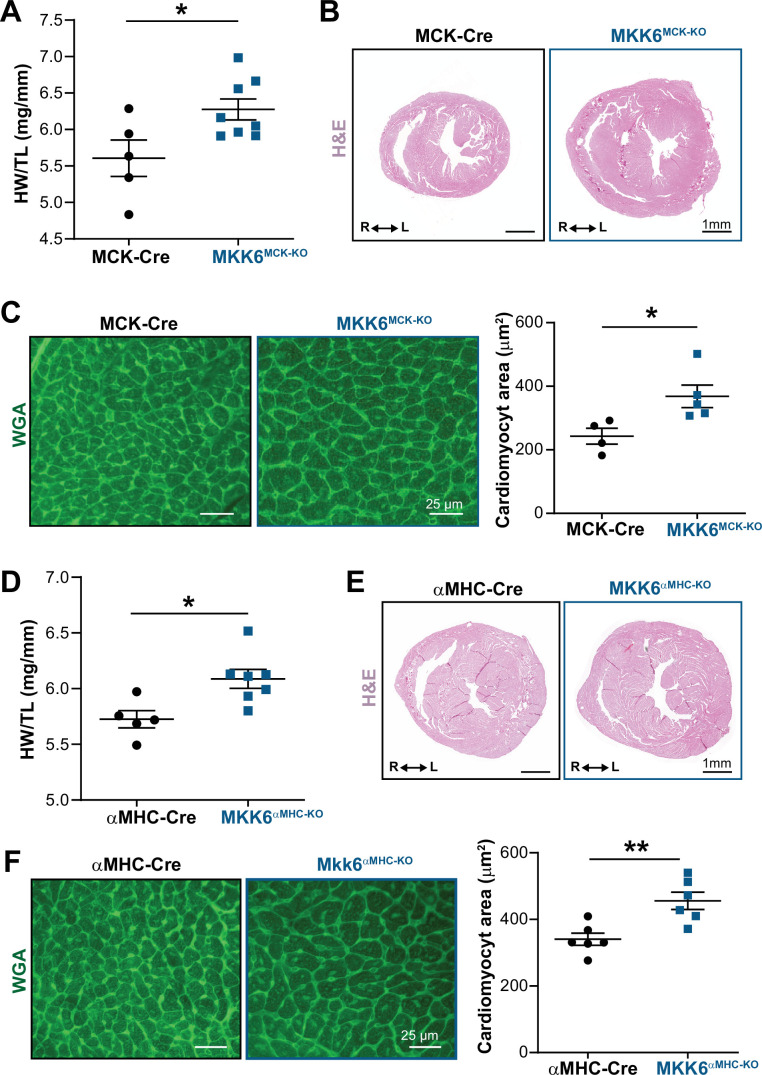
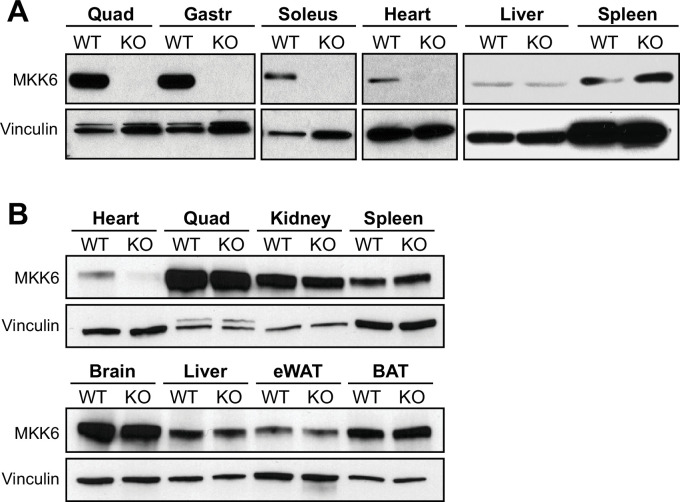
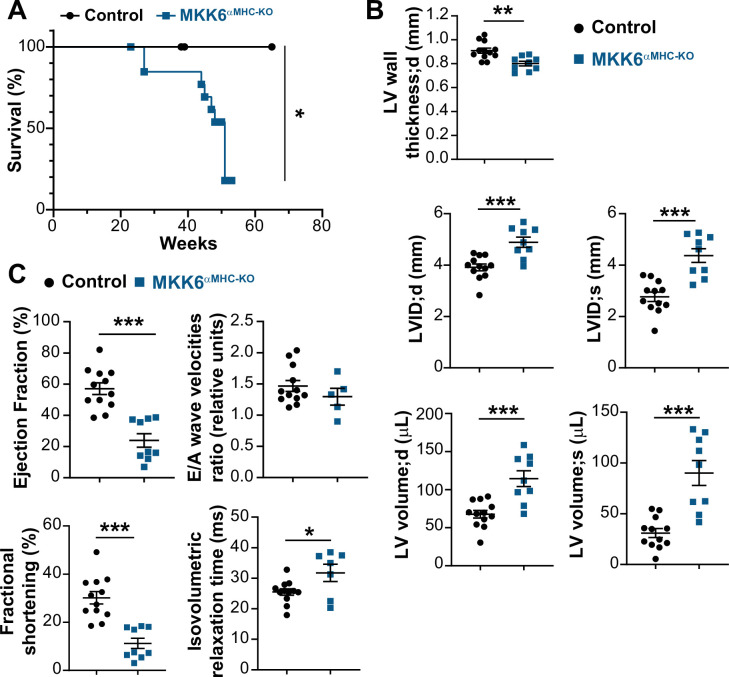
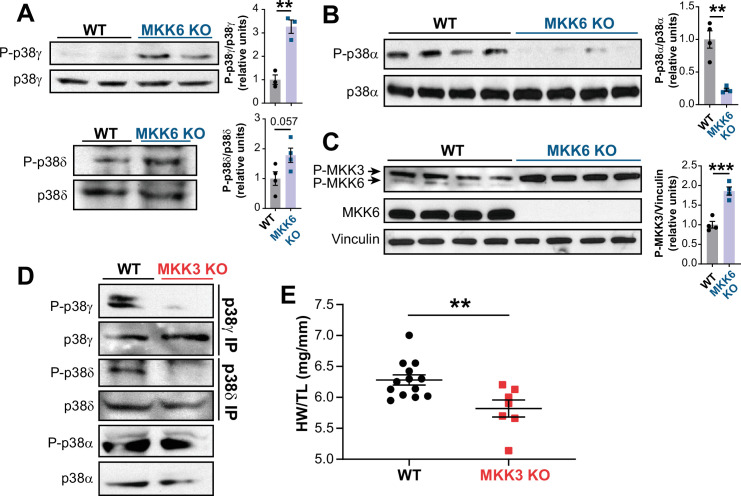
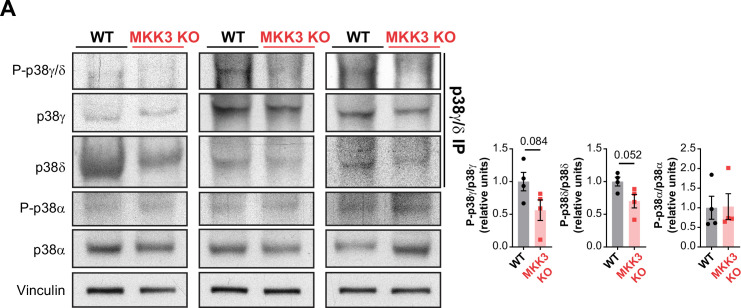
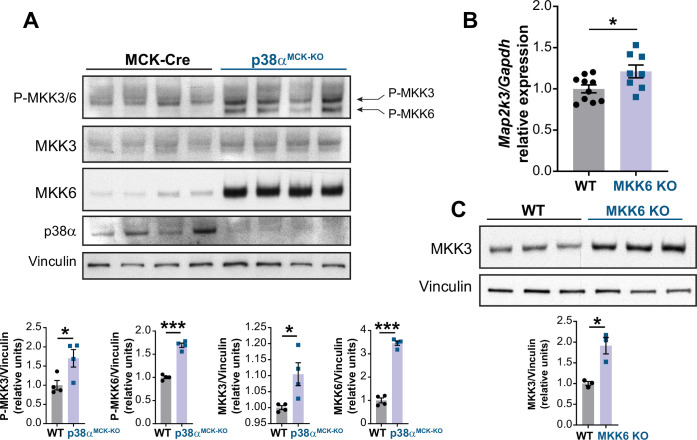
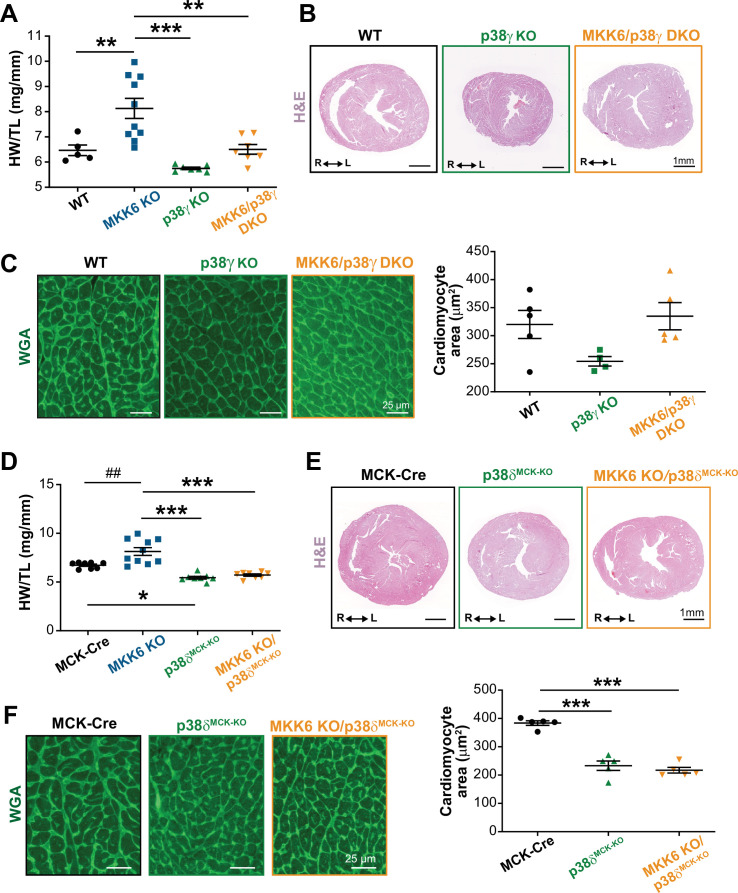
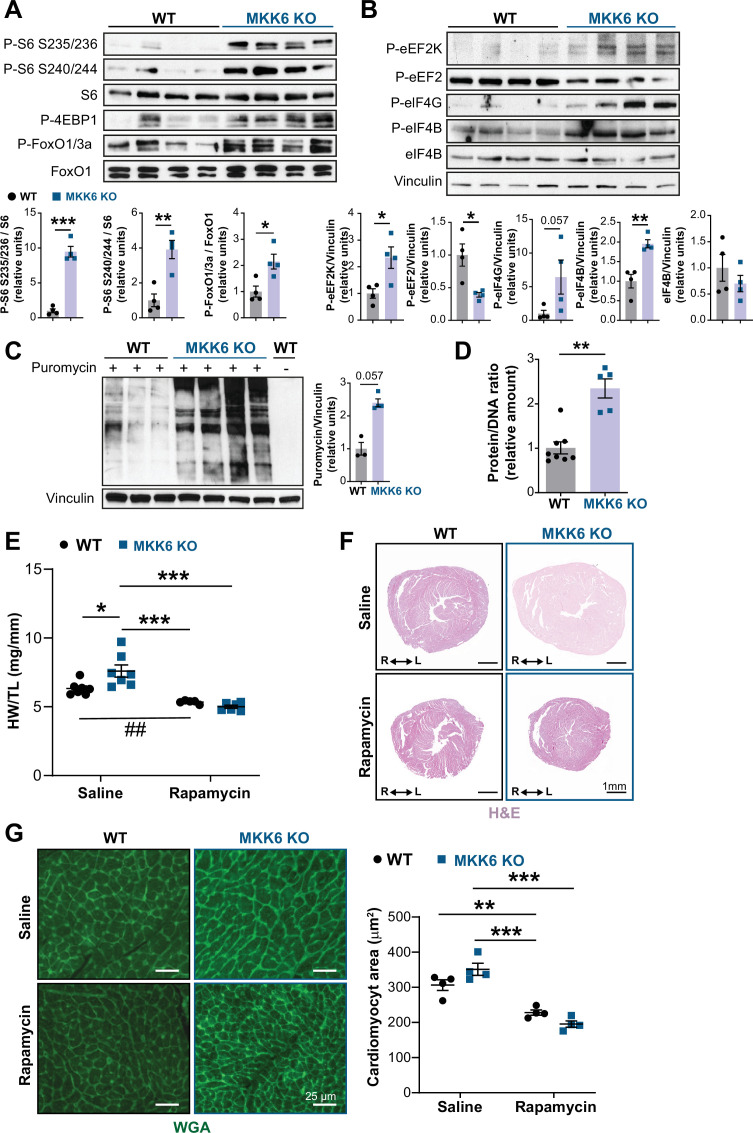
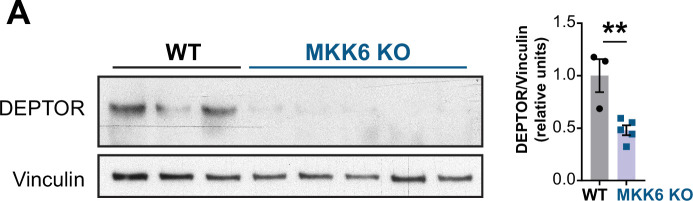
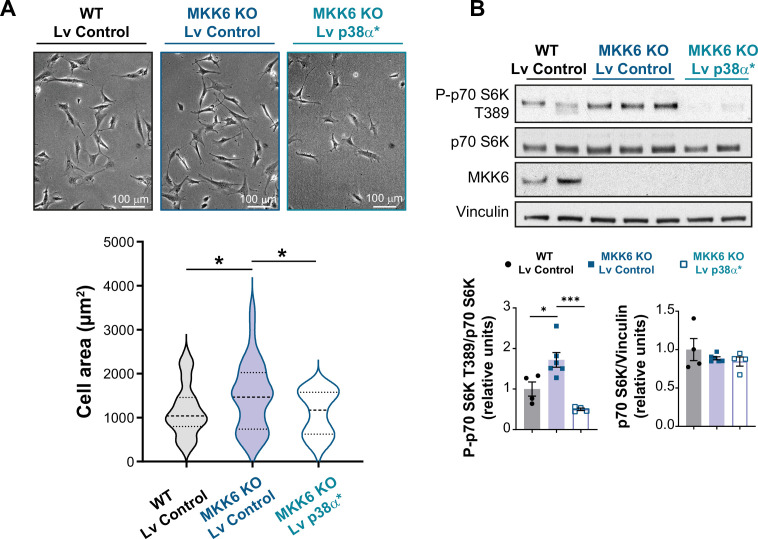
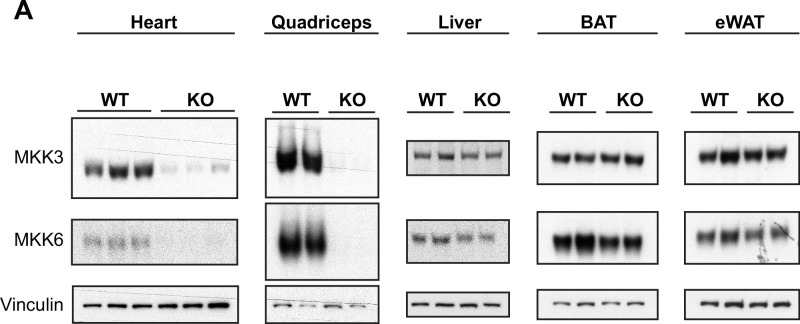
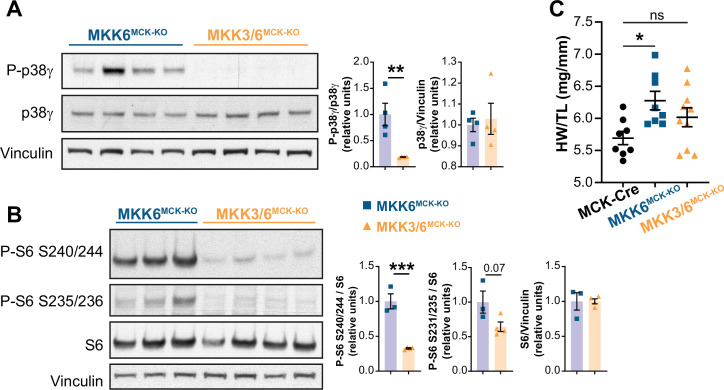
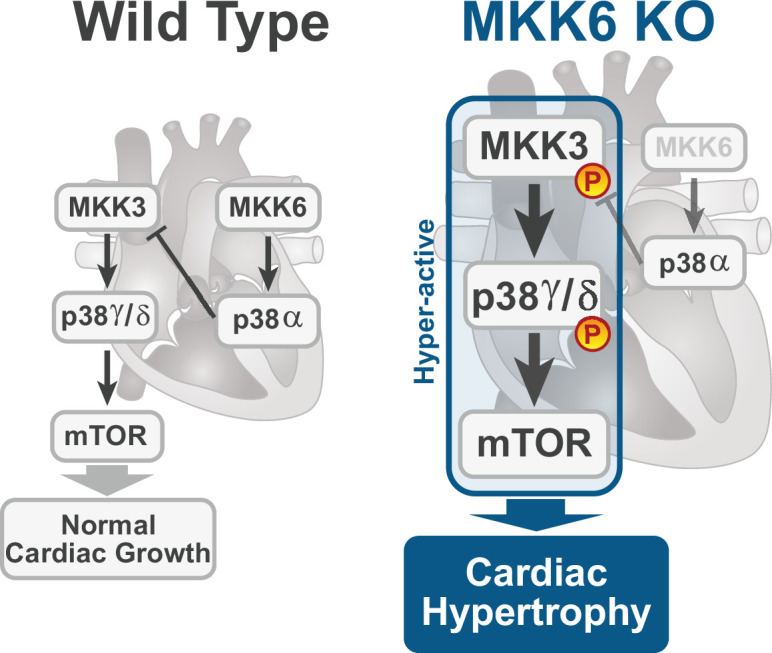
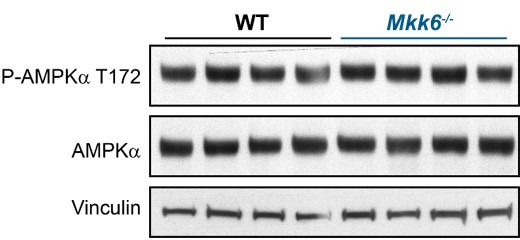
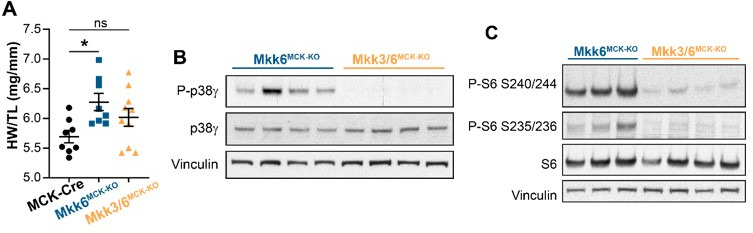
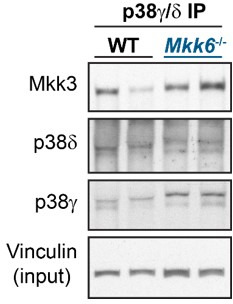
Stress-activated p38 kinases control a plethora of functions, and their dysregulation has been linked to the development of steatosis, obesity, immune disorders, and cancer. Therefore, they have been identified as potential targets for novel therapeutic strategies. There are four p38 family members (p38α, p38β, p38γ, and p38δ) that are activated by MKK3 and MKK6. Here, we demonstrate that lack of MKK6 reduces the lifespan in mice. Longitudinal study of cardiac function in MKK6 KO mice showed that young mice develop cardiac hypertrophy which progresses to cardiac dilatation and fibrosis with age. Mechanistically, lack of MKK6 blunts p38α activation while causing MKK3-p38γ/δ hyperphosphorylation and increased mammalian target of rapamycin (mTOR) signaling, resulting in cardiac hypertrophy. Cardiac hypertrophy in MKK6 KO mice is reverted by knocking out either p38γ or p38δ or by inhibiting the mTOR pathway with rapamycin. In conclusion, we have identified a key role for the MKK3/6-p38γ/δ pathway in the development of cardiac hypertrophy, which has important implications for the clinical use of p38α inhibitors in the long-term treatment since they might result in cardiotoxicity.
应激激活的 p38 激酶控制着大量的功能,其失调与脂肪变性、肥胖、免疫紊乱和癌症的发展有关。因此,它们被认为是新的治疗策略的潜在靶点。有四个 p38 家族成员(p38α、p38β、p38γ 和 p38δ)被 MKK3 和 MKK6 激活。在这里,我们证明缺乏 MKK6 会缩短小鼠的寿命。对 MKK6 KO 小鼠心脏功能的纵向研究表明,年轻小鼠会发生心脏肥大,随着年龄的增长,会进展为心脏扩张和纤维化。从机制上讲,缺乏 MKK6 会抑制 p38α 的激活,同时导致 MKK3-p38γ/δ 的过度磷酸化和哺乳动物雷帕霉素靶蛋白(mTOR)信号的增加,导致心脏肥大。用 rapamycin 敲除 p38γ 或 p38δ 或抑制 mTOR 通路可使 MKK6 KO 小鼠的心脏肥大逆转。总之,我们确定了 MKK3/6-p38γ/δ 通路在心脏肥大发展中的关键作用,这对 p38α 抑制剂在长期治疗中的临床应用具有重要意义,因为它们可能导致心脏毒性。