Department of Ecophysiology, Max Planck Institute for Terrestrial Microbiologygrid.419554.8, Marburg, Germany.
Core Facility for Mass Spectrometry & Proteomics, Max Planck Institute for Terrestrial Microbiologygrid.419554.8, Marburg, Germany.
mBio. 2022 Oct 26;13(5):e0203222. doi: 10.1128/mbio.02032-22. Epub 2022 Aug 16.
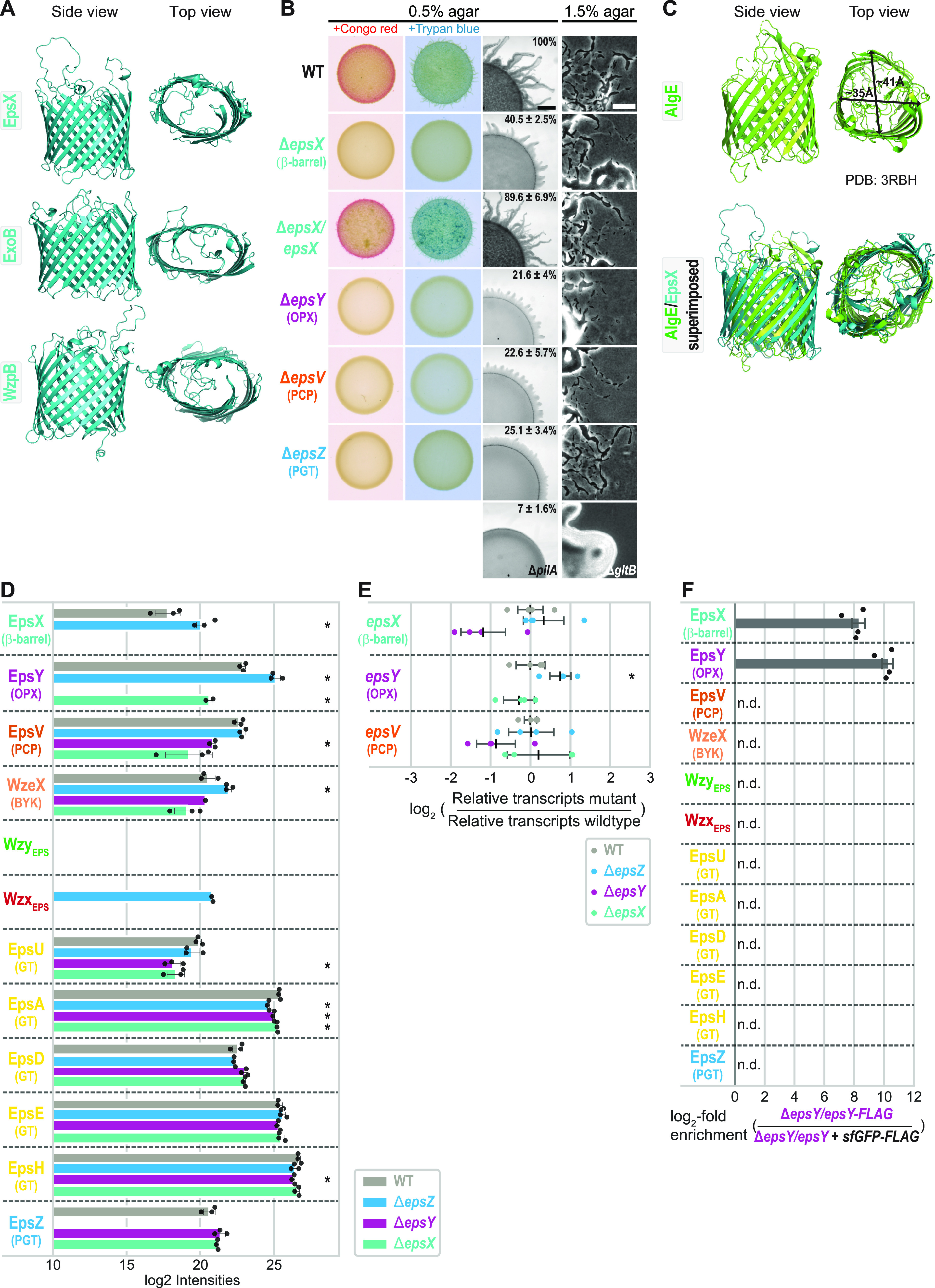
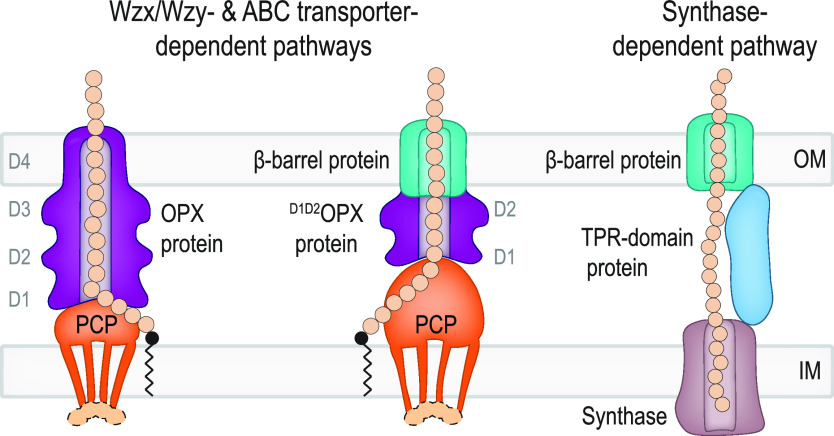
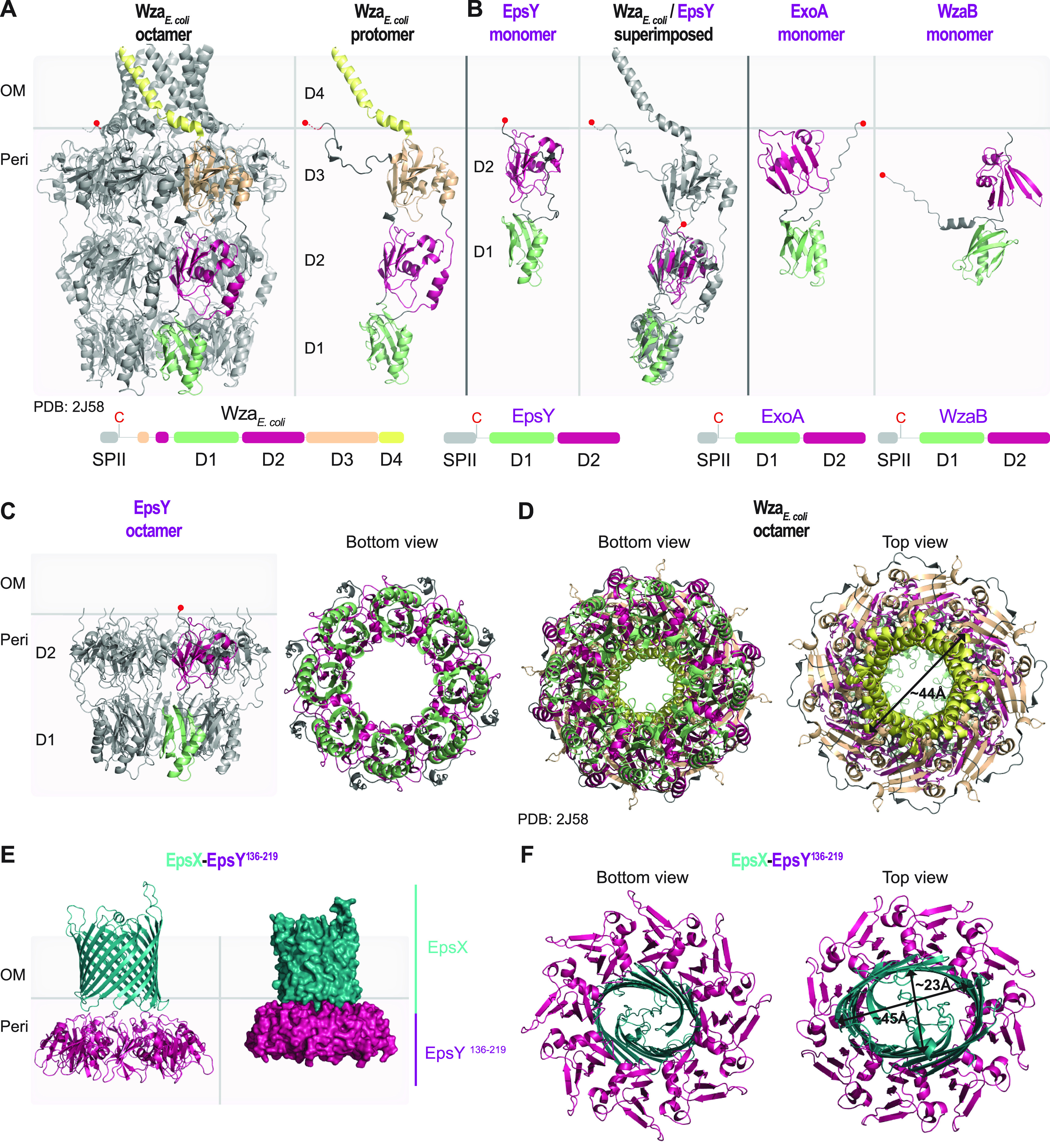
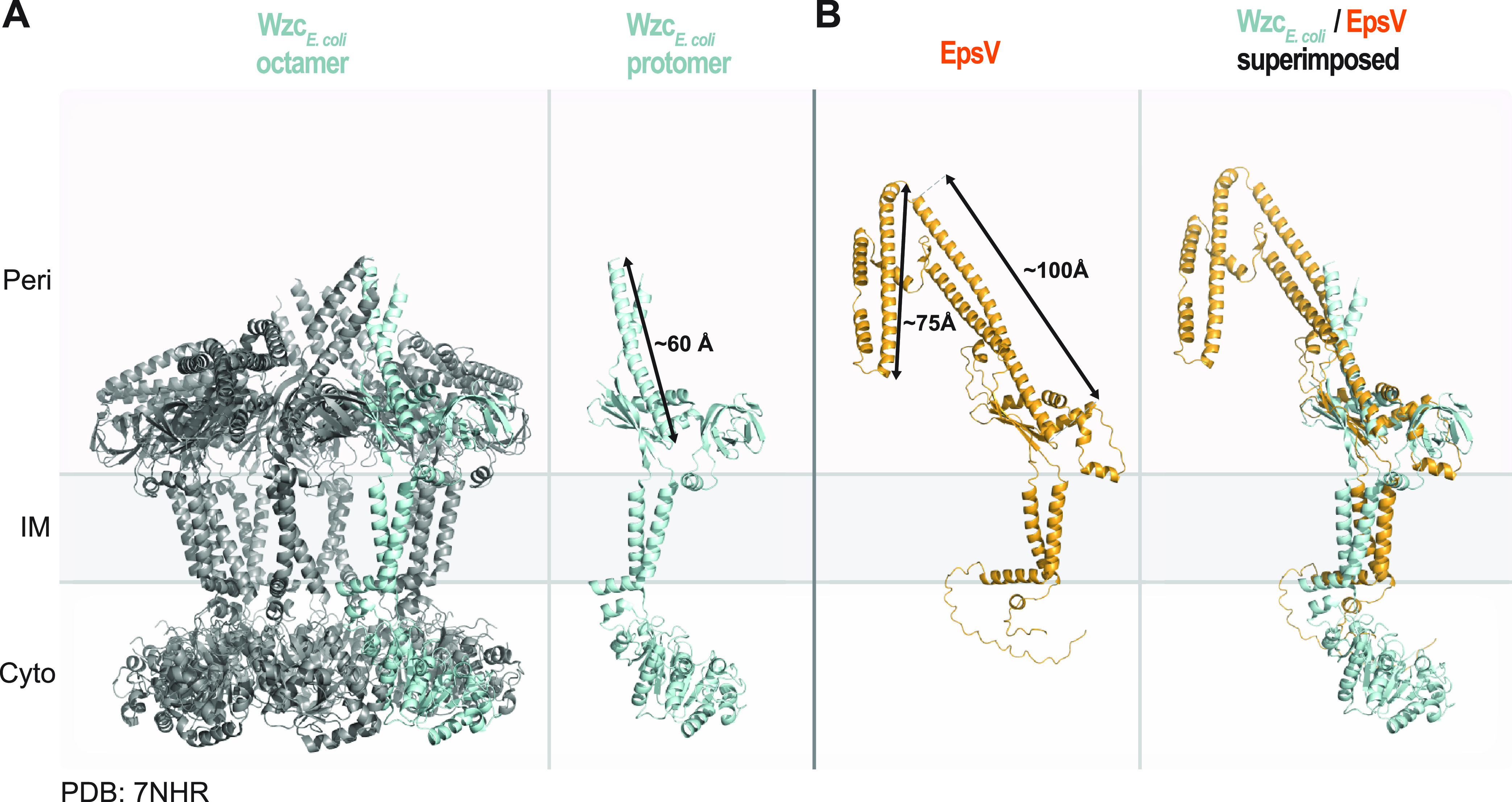
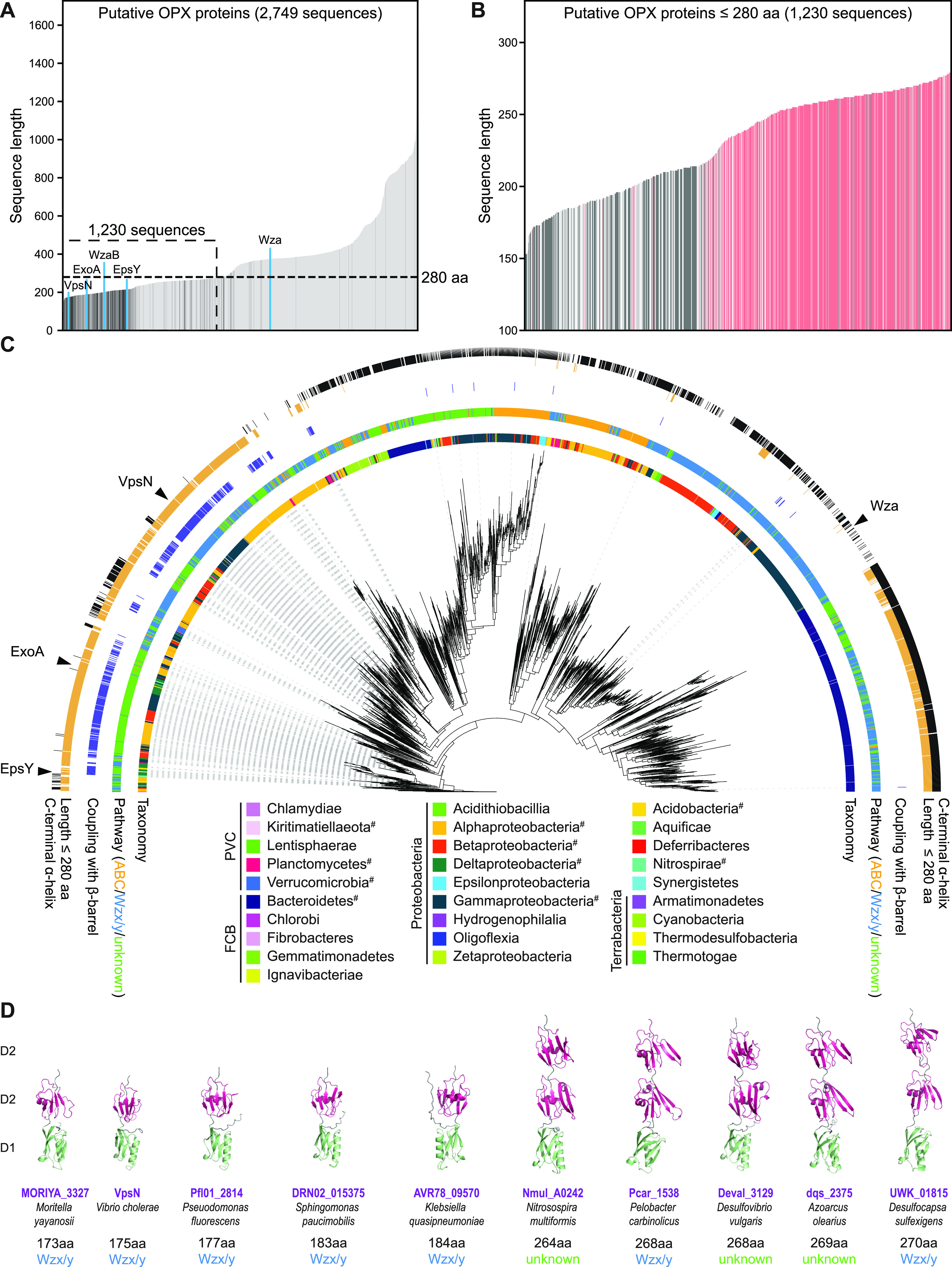
In Gram-negative bacteria, secreted polysaccharides have multiple critical functions. In Wzx/Wzy- and ABC transporter-dependent pathways, an outer membrane (OM) polysaccharide export (OPX) type translocon exports the polysaccharide across the OM. The paradigm OPX protein Wza of Escherichia coli is an octamer in which the eight C-terminal domains form an α-helical OM pore and the eight copies of the three N-terminal domains (D1 to D3) form a periplasmic cavity. In synthase-dependent pathways, the OM translocon is a 16- to 18-stranded β-barrel protein. In Myxococcus xanthus, the secreted polysaccharide EPS (exopolysaccharide) is synthesized in a Wzx/Wzy-dependent pathway. Here, using experiments, phylogenomics, and computational structural biology, we identify and characterize EpsX as an OM 18-stranded β-barrel protein important for EPS synthesis and identify AlgE, a β-barrel translocon of a synthase-dependent pathway, as its closest structural homolog. We also find that EpsY, the OPX protein of the EPS pathway, consists only of the periplasmic D1 and D2 domains and completely lacks the domain for spanning the OM (herein termed a OPX protein). , EpsX and EpsY mutually stabilize each other and interact in pulldown experiments supporting their direct interaction. Based on these observations, we propose that EpsY and EpsX make up and represent a third type of translocon for polysaccharide export across the OM. Specifically, in this composite translocon, EpsX functions as the OM-spanning β-barrel translocon together with the periplasmic OPX protein EpsY. Based on computational genomics, similar composite systems are widespread in Gram-negative bacteria. Bacteria secrete a wide variety of polysaccharides that have critical functions in, e.g., fitness, surface colonization, and biofilm formation and in beneficial and pathogenic human-, animal-, and plant-microbe interactions. In Gram-negative bacteria, export of these chemically diverse polysaccharides across the outer membrane depends on two known translocons, i.e., an outer membrane OPX protein in Wzx/Wzy- and ABC transporter-dependent pathways and an outer membrane 16- to 18-stranded β-barrel protein in synthase-dependent pathways. Here, using a combination of experiments in Myxococcus xanthus, phylogenomics, and computational structural biology, we provide evidence supporting that a third type of translocon can export polysaccharides across the outer membrane. Specifically, in this translocon, an outer membrane-spanning β-barrel protein functions together with an entirely periplasmic OPX protein that completely lacks the domain for spanning the OM. Computational genomics support that similar composite systems are widespread in Gram-negative bacteria.
在革兰氏阴性菌中,分泌的多糖具有多种关键功能。在 Wzx/Wzy 和 ABC 转运蛋白依赖途径中,一种外膜 (OM) 多糖输出 (OPX) 型转运蛋白将多糖跨 OM 输出。大肠杆菌的典范 OPX 蛋白 Wza 是一个八聚体,其中八个 C 末端结构域形成一个 α 螺旋 OM 孔,而八个 N 末端结构域 (D1 至 D3) 形成周质腔。在合成酶依赖途径中,OM 转运蛋白是一个 16 到 18 股的 β 桶蛋白。在粘细菌中,分泌的多糖 EPS(胞外多糖)是在 Wzx/Wzy 依赖途径中合成的。在这里,我们使用实验、系统发育基因组学和计算结构生物学,鉴定并表征 EpsX 作为一种 OM 18 股 β 桶蛋白,对 EPS 合成很重要,并鉴定 AlgE,一种合成酶依赖途径的 β 桶转运蛋白,作为其最接近的结构同源物。我们还发现,EPS 途径的 OPX 蛋白 EpsY 仅由周质腔中的 D1 和 D2 结构域组成,完全缺乏跨越 OM 的结构域(在此称为 OPX 蛋白)。EpsX 和 EpsY 相互稳定,并在下拉实验中相互作用,支持它们的直接相互作用。基于这些观察结果,我们提出 EpsY 和 EpsX 构成并代表了一种用于跨 OM 输出多糖的第三种转运蛋白。具体来说,在这种复合转运蛋白中,EpsX 与周质腔中的 OPX 蛋白 EpsY 一起作为 OM 跨越的 β 桶转运蛋白发挥作用。基于计算基因组学,类似的复合系统在革兰氏阴性菌中广泛存在。细菌分泌多种具有关键功能的多糖,例如在适应度、表面定植和生物膜形成以及有益和致病的人类、动物和植物微生物相互作用中。在革兰氏阴性菌中,这些化学多样化的多糖跨外膜的输出取决于两种已知的转运蛋白,即在 Wzx/Wzy 和 ABC 转运蛋白依赖途径中的外膜 OPX 蛋白和在合成酶依赖途径中的外膜 16-18 股 β 桶蛋白。在这里,我们使用粘细菌中的一系列实验、系统发育基因组学和计算结构生物学,提供了支持第三种转运蛋白可以跨外膜输出多糖的证据。具体来说,在这种转运蛋白中,一种外膜跨越的 β 桶蛋白与完全缺乏跨 OM 结构域的整个周质腔 OPX 蛋白一起发挥作用。计算基因组学支持类似的复合系统在革兰氏阴性菌中广泛存在。