Frasch Wayne D, Bukhari Zain A, Yanagisawa Seiga
School of Life Sciences, Arizona State University, Tempe, AZ, United States.
Front Microbiol. 2022 Aug 23;13:965620. doi: 10.3389/fmicb.2022.965620. eCollection 2022.
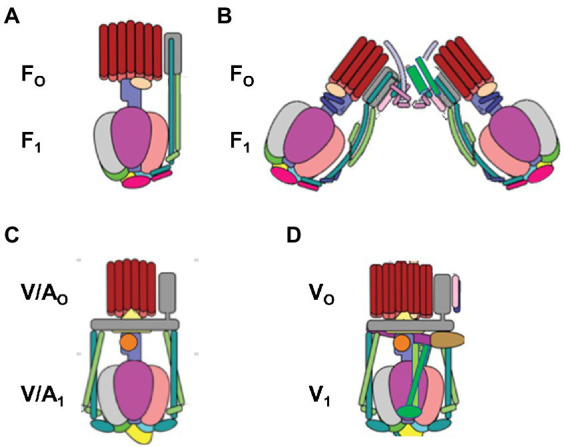
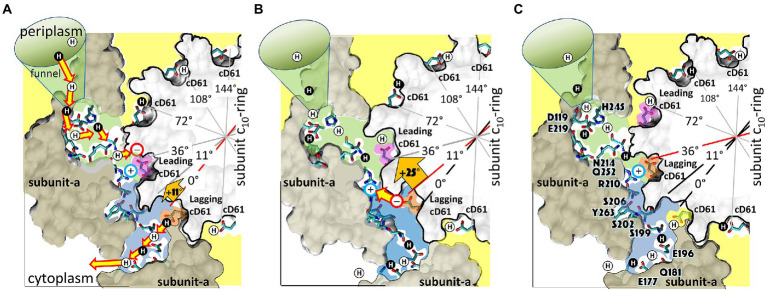
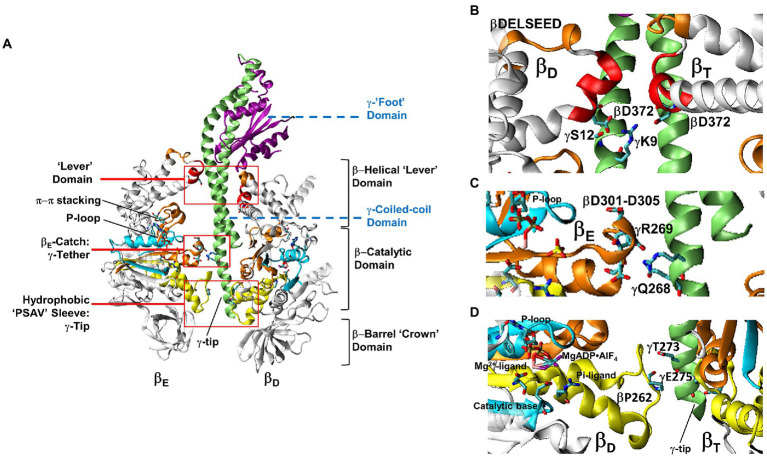
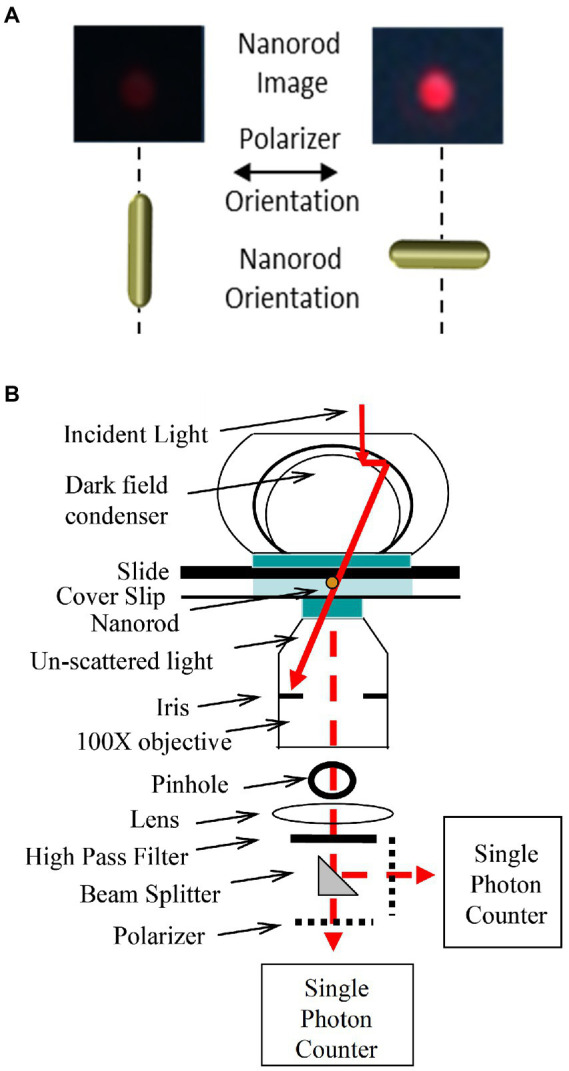
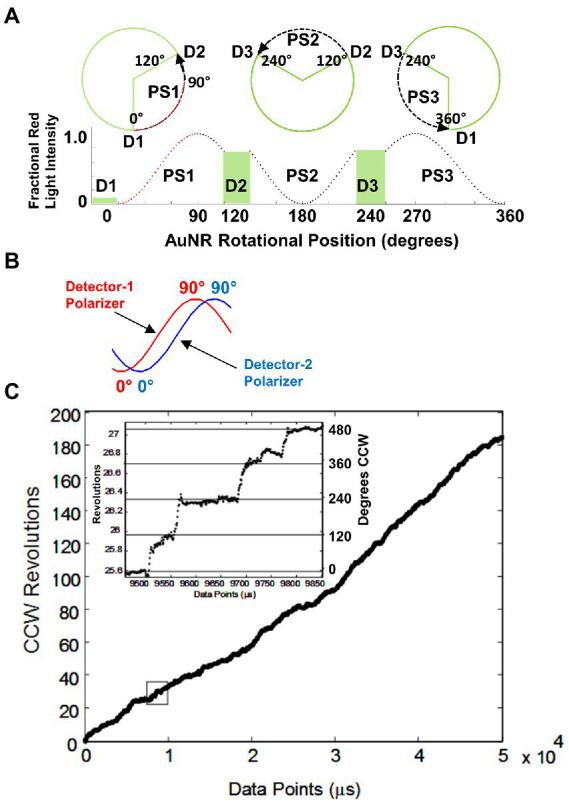
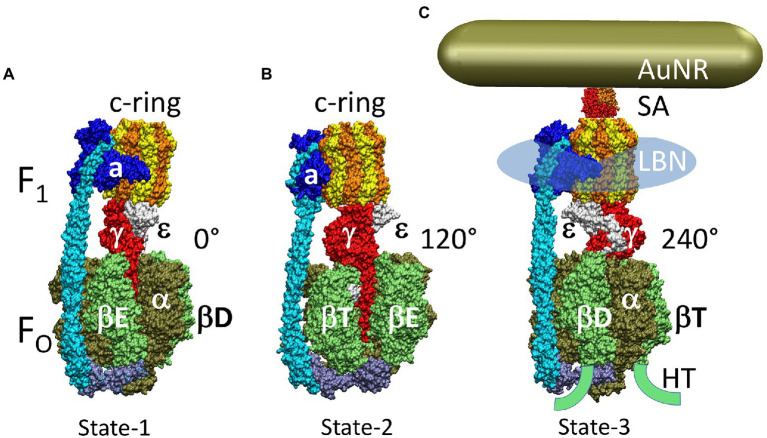
The F-ATP synthase, consisting of F and F motors connected by a central rotor and the stators, is the enzyme responsible for synthesizing the majority of ATP in all organisms. The F (αβ) ring stator contains three catalytic sites. Single-molecule F rotation studies revealed that ATP hydrolysis at each catalytic site (0°) precedes a power-stroke that rotates subunit-γ 120° with angular velocities that vary with rotational position. Catalytic site conformations vary relative to subunit-γ position (β, empty; β, ADP bound; β, ATP-bound). During a power stroke, β binds ATP (0°-60°) and β releases ADP (60°-120°). Årrhenius analysis of the power stroke revealed that elastic energy powers rotation unwinding the γ-subunit coiled-coil. Energy from ATP binding at 34° closes β upon subunit-γ to drive rotation to 120° and forcing the subunit-γ to exchange its tether from β to β, which changes catalytic site conformations. In FF, the membrane-bound F complex contains a ring of c-subunits that is attached to subunit-γ. This c-ring rotates relative to the subunit-a stator in response to transmembrane proton flow driven by a pH gradient, which drives subunit-γ rotation in the opposite direction to force ATP synthesis in F. Single-molecule studies of FF embedded in lipid bilayer nanodisks showed that the c-ring transiently stopped F-ATPase-driven rotation every 36° (at each c-subunit in the c-ring of FF) and was able to rotate 11° in the direction of ATP synthesis. Protonation and deprotonation of the conserved carboxyl group on each c-subunit is facilitated by separate groups of subunit-a residues, which were determined to have different pKa's. Mutations of any of any residue from either group changed both pKa values, which changed the occurrence of the 11° rotation proportionately. This supports a Grotthuss mechanism for proton translocation and indicates that proton translocation occurs during the 11° steps. This is consistent with a mechanism in which each 36° of rotation the c-ring during ATP synthesis involves a proton translocation-dependent 11° rotation of the c-ring, followed by a 25° rotation driven by electrostatic interaction of the negatively charged unprotonated carboxyl group to the positively charged essential arginine in subunit-a.
F-ATP合酶由通过中央转子和定子连接的F和F马达组成,是负责在所有生物体中合成大部分ATP的酶。F(αβ)环定子包含三个催化位点。单分子F旋转研究表明,每个催化位点(0°)处的ATP水解先于一个动力冲程,该动力冲程使亚基γ旋转120°,其角速度随旋转位置而变化。催化位点构象相对于亚基γ位置有所不同(β,空;β,结合ADP;β,结合ATP)。在动力冲程期间,β结合ATP(0°-60°),β释放ADP(60°-120°)。对动力冲程的阿累尼乌斯分析表明,弹性能量为旋转提供动力,解开γ亚基的卷曲螺旋。34°时ATP结合产生的能量使β在亚基γ上关闭,以驱动旋转至120°,并迫使亚基γ将其系链从β交换到β,这改变了催化位点构象。在FF中,膜结合的F复合物包含一个与亚基γ相连的c亚基环。该c环响应由pH梯度驱动的跨膜质子流相对于亚基a定子旋转,这驱动亚基γ向相反方向旋转以迫使F中合成ATP。对嵌入脂质双层纳米盘中的FF进行的单分子研究表明,c环每36°(在FF的c环中的每个c亚基处)短暂停止F-ATP酶驱动的旋转,并能够沿ATP合成方向旋转11°。每个c亚基上保守羧基的质子化和去质子化由亚基a残基的不同组促进,这些残基被确定具有不同的pKa值。来自任何一组的任何残基的突变都会改变两个pKa值,这相应地改变了11°旋转的发生率。这支持了质子转运的Grotthuss机制,并表明质子转运发生在11°步骤期间。这与一种机制一致,即在ATP合成过程中,c环每旋转36°涉及c环质子转运依赖性的11°旋转,随后是由带负电荷的未质子化羧基与亚基a中带正电荷的必需精氨酸的静电相互作用驱动的25°旋转。