Janelia Research Campus, Ashburn, United States.
Physiology Institute and Millennium Nucleus of Ion Channel-Associated Diseases, Universidad Austral de Chile, Valdivia, Chile.
Elife. 2023 Jan 25;12:e84477. doi: 10.7554/eLife.84477.
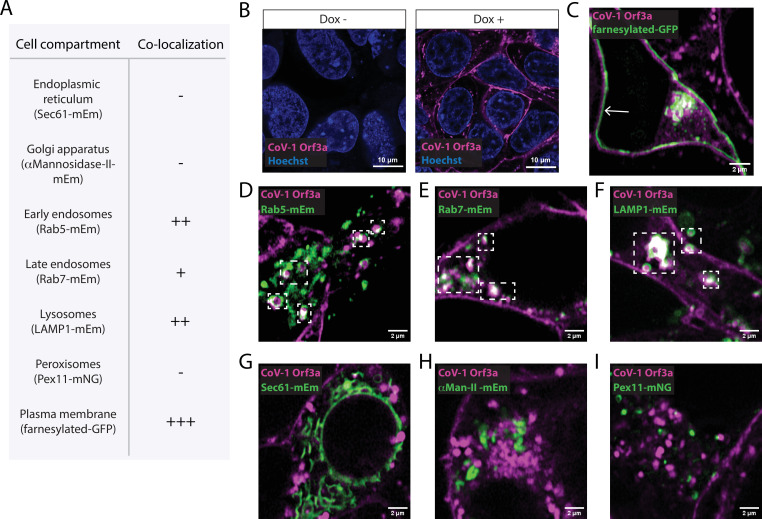
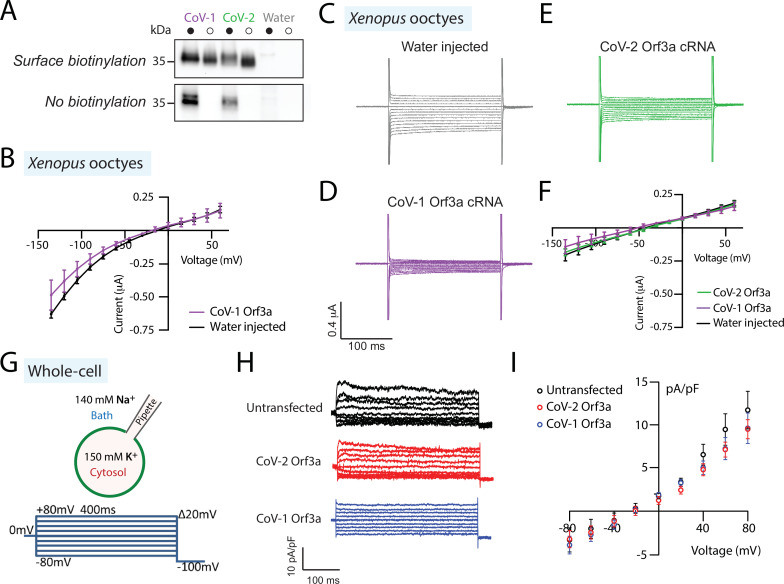
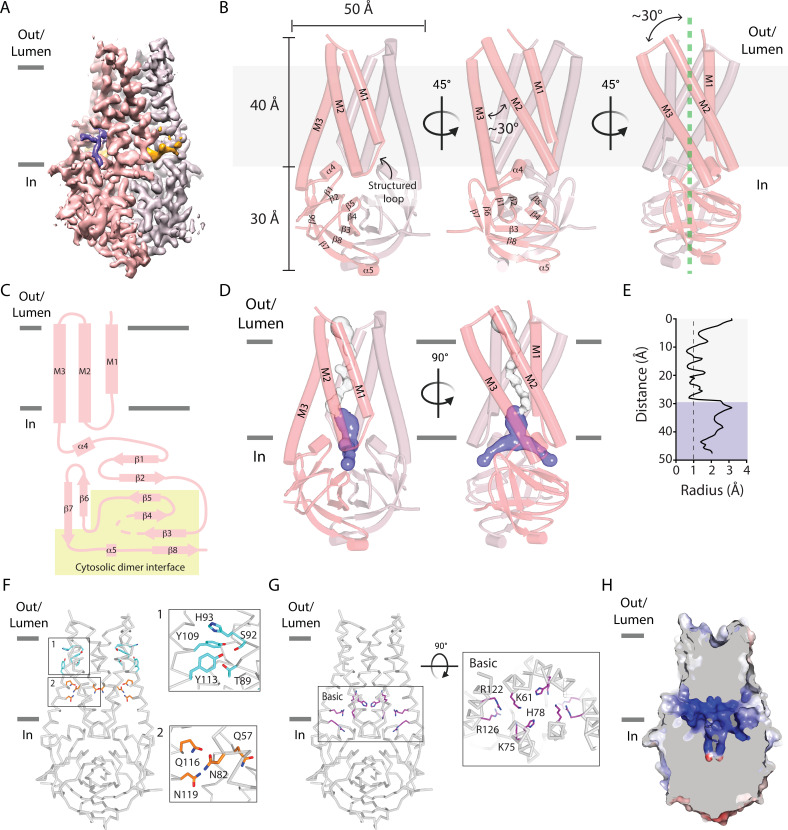
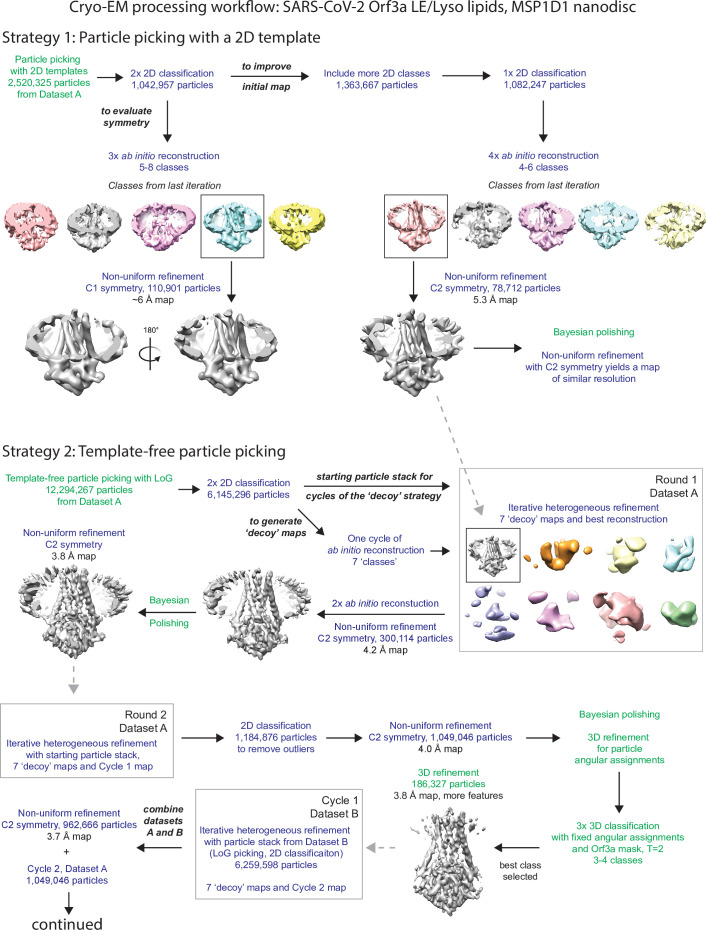
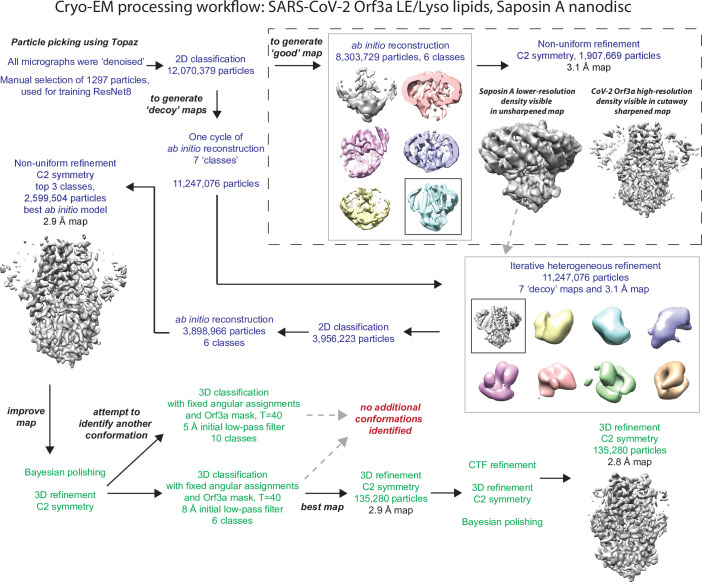
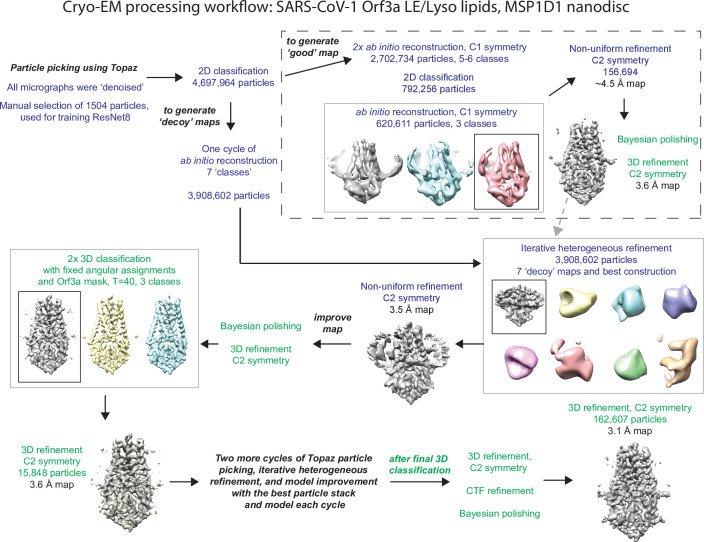
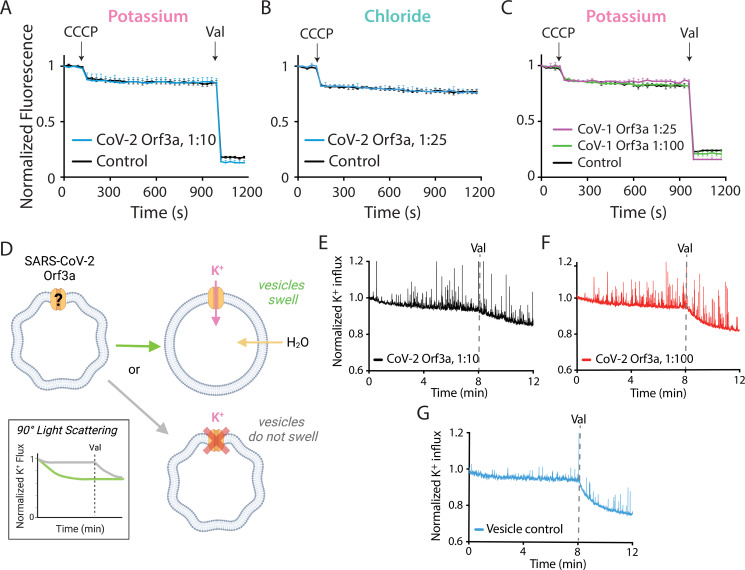
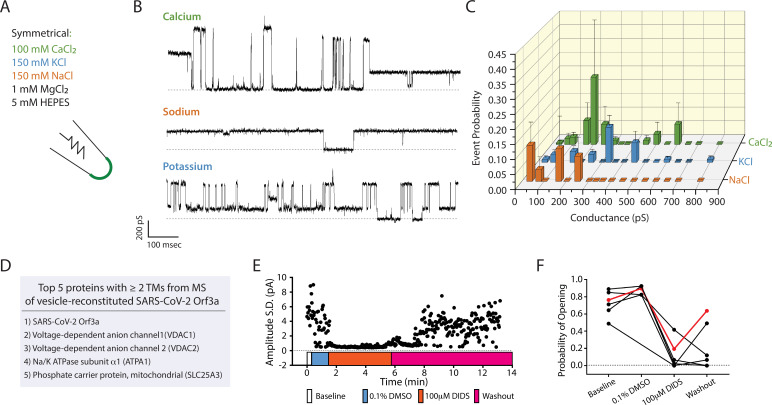
The severe acute respiratory syndrome associated coronavirus 2 (SARS-CoV-2) and SARS-CoV-1 accessory protein Orf3a colocalizes with markers of the plasma membrane, endocytic pathway, and Golgi apparatus. Some reports have led to annotation of both Orf3a proteins as viroporins. Here, we show that neither SARS-CoV-2 nor SARS-CoV-1 Orf3a form functional ion conducting pores and that the conductances measured are common contaminants in overexpression and with high levels of protein in reconstitution studies. Cryo-EM structures of both SARS-CoV-2 and SARS-CoV-1 Orf3a display a narrow constriction and the presence of a positively charged aqueous vestibule, which would not favor cation permeation. We observe enrichment of the late endosomal marker Rab7 upon SARS-CoV-2 Orf3a overexpression, and co-immunoprecipitation with VPS39. Interestingly, SARS-CoV-1 Orf3a does not cause the same cellular phenotype as SARS-CoV-2 Orf3a and does not interact with VPS39. To explain this difference, we find that a divergent, unstructured loop of SARS-CoV-2 Orf3a facilitates its binding with VPS39, a HOPS complex tethering protein involved in late endosome and autophagosome fusion with lysosomes. We suggest that the added loop enhances SARS-CoV-2 Orf3a's ability to co-opt host cellular trafficking mechanisms for viral exit or host immune evasion.
严重急性呼吸系统综合症相关冠状病毒 2(SARS-CoV-2)和 SARS-CoV-1 辅助蛋白 Orf3a 与质膜、内吞途径和高尔基体的标志物共定位。一些报道导致两个 Orf3a 蛋白都被注释为病毒孔蛋白。在这里,我们表明,SARS-CoV-2 和 SARS-CoV-1 的 Orf3a 都不形成功能性离子通道,并且在过表达和高蛋白质重建研究中测量的电导都是常见的污染物。SARS-CoV-2 和 SARS-CoV-1 Orf3a 的冷冻电镜结构显示出狭窄的收缩和带正电荷的水前庭的存在,这不利于阳离子渗透。我们观察到 SARS-CoV-2 Orf3a 过表达时晚期内体标志物 Rab7 的富集,以及与 VPS39 的共免疫沉淀。有趣的是,SARS-CoV-1 Orf3a 不会引起与 SARS-CoV-2 Orf3a 相同的细胞表型,也不会与 VPS39 相互作用。为了解释这种差异,我们发现 SARS-CoV-2 Orf3a 的一个发散的、无结构的环促进了它与 VPS39 的结合,VPS39 是一种参与晚期内体和自噬体与溶酶体融合的 HOPS 复合物连接蛋白。我们认为,附加环增强了 SARS-CoV-2 Orf3a 劫持宿主细胞运输机制进行病毒释放或宿主免疫逃避的能力。