School of Biological Sciences, Seoul National University, Seoul, Republic of Korea.
Research Center for Plant Plasticity, Seoul National University, Seoul, Republic of Korea.
Elife. 2023 Feb 1;12:e84594. doi: 10.7554/eLife.84594.
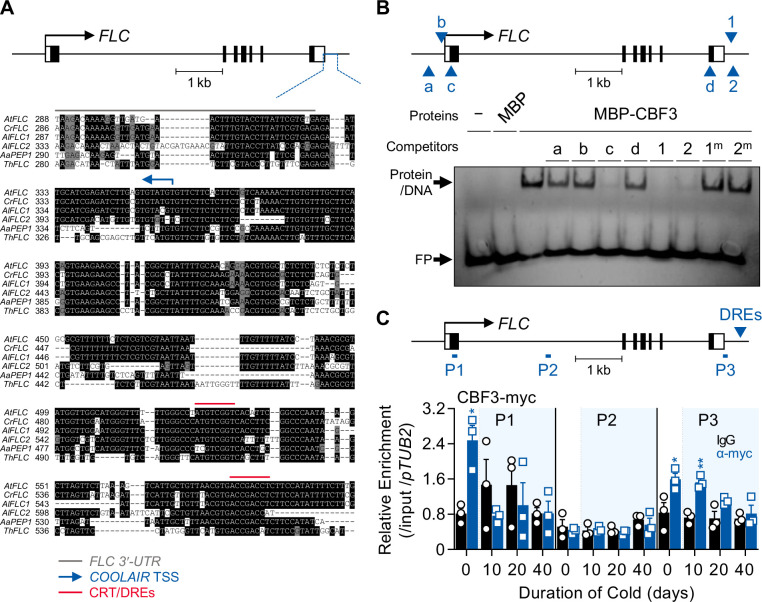
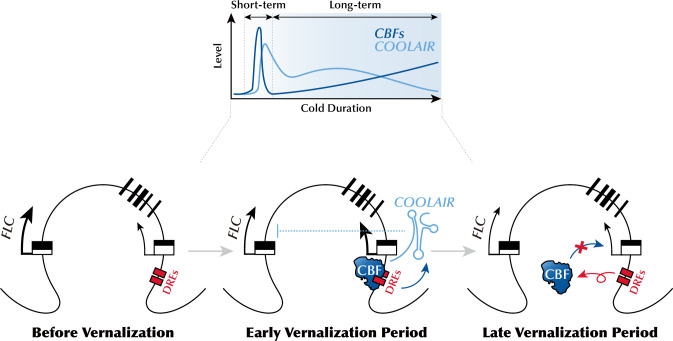
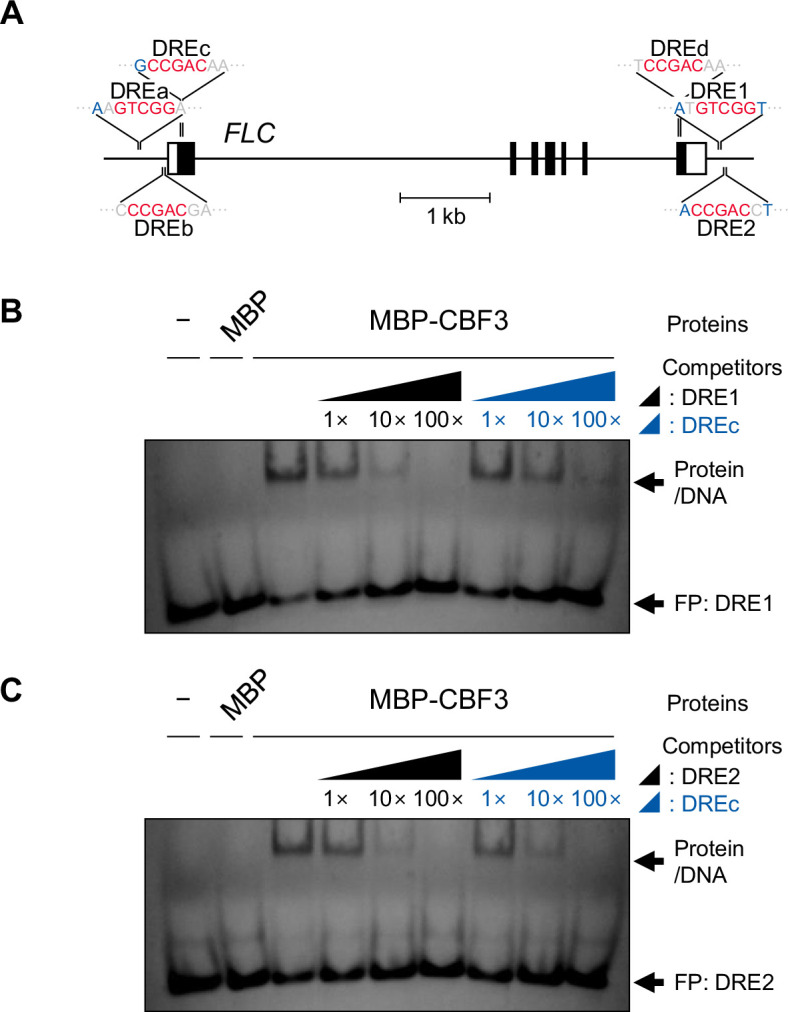
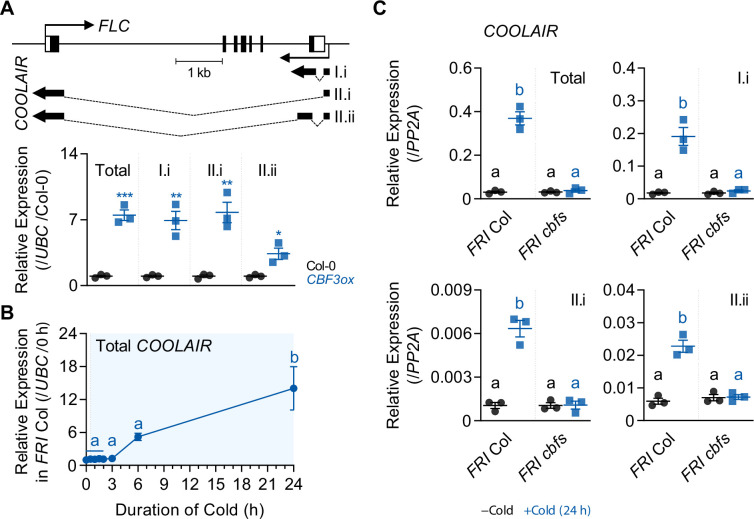
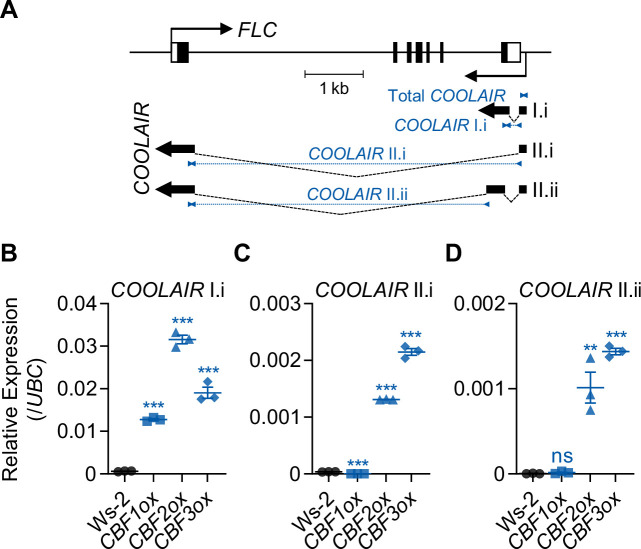
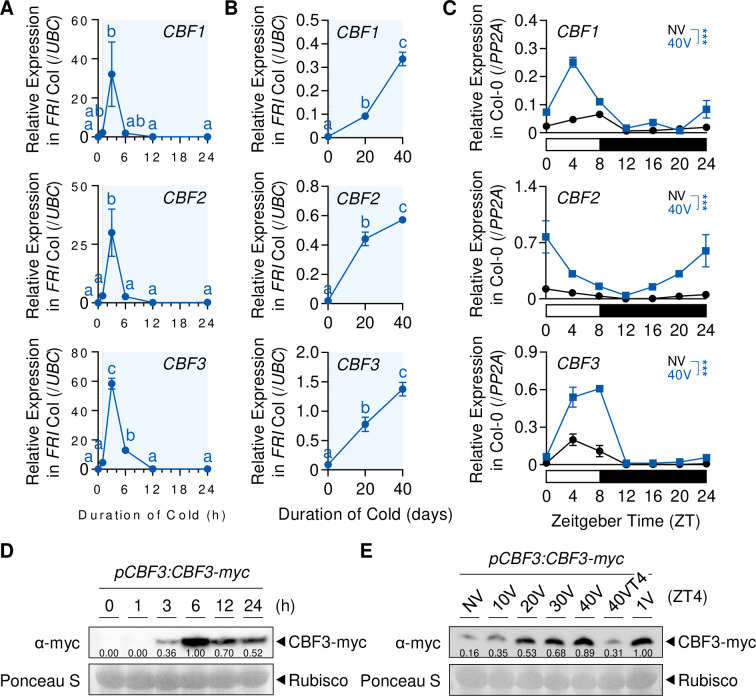
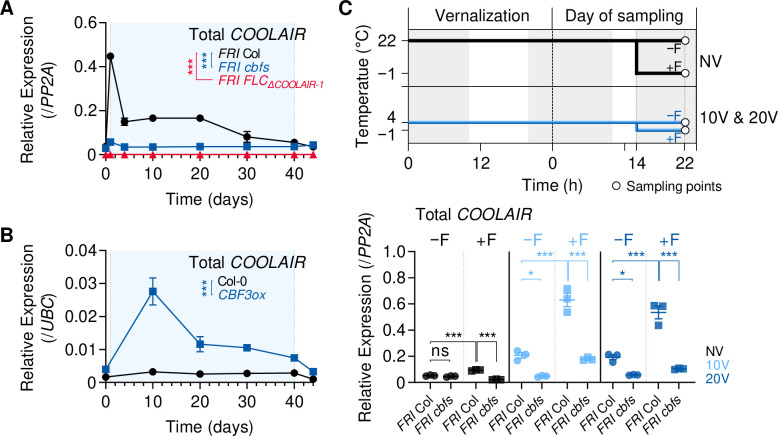
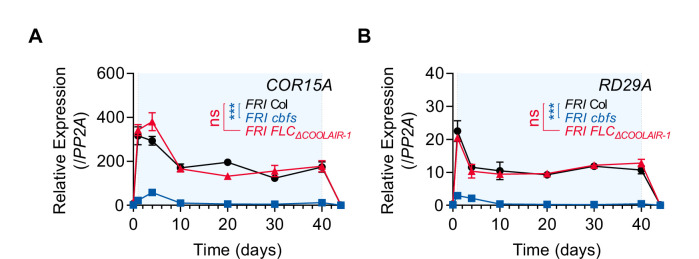
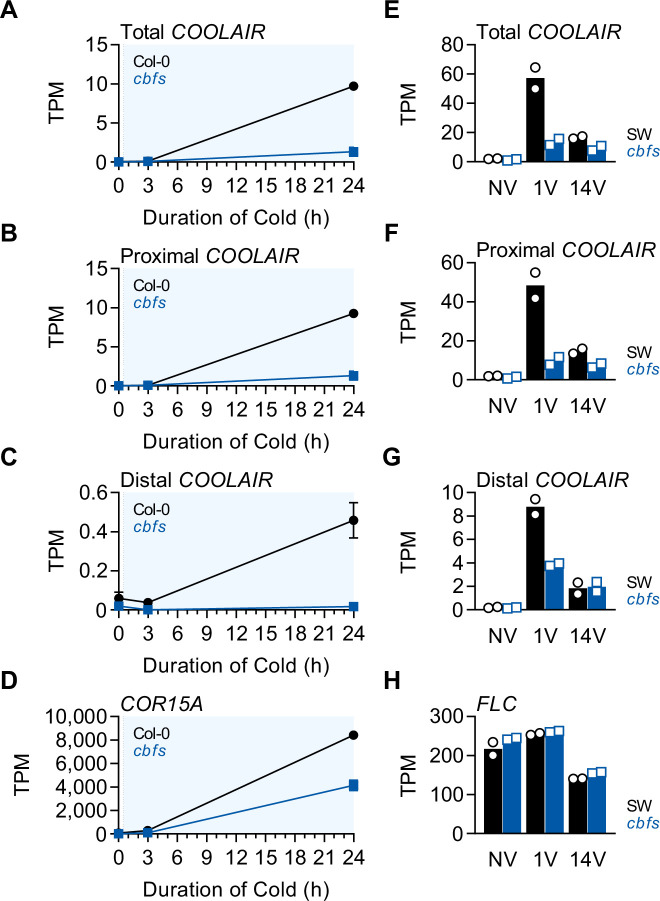
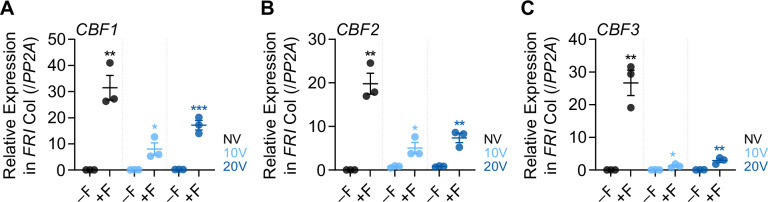
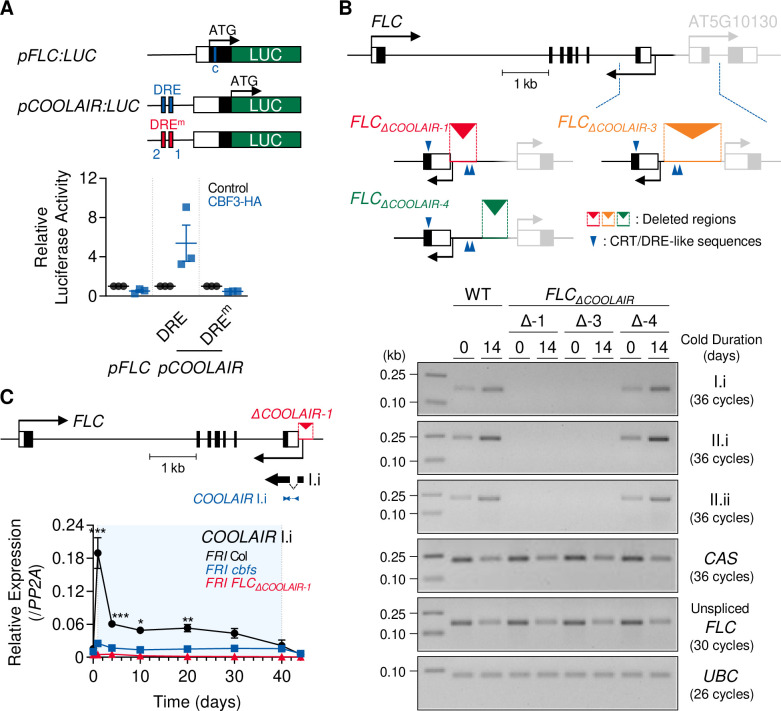
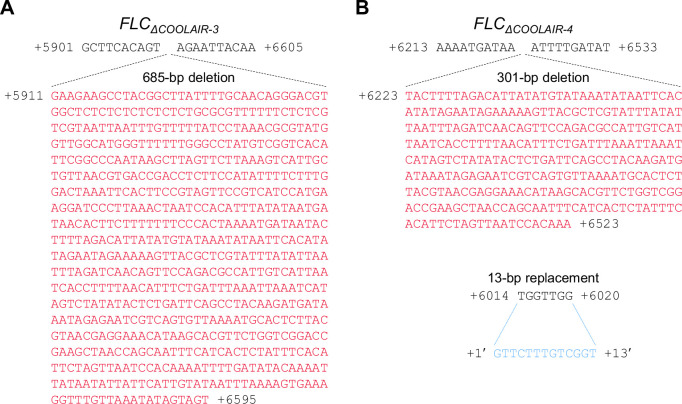
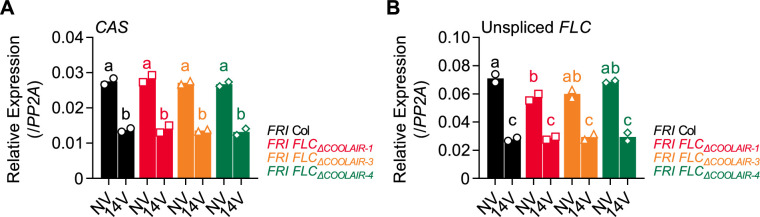
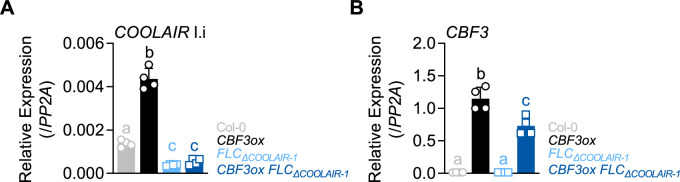
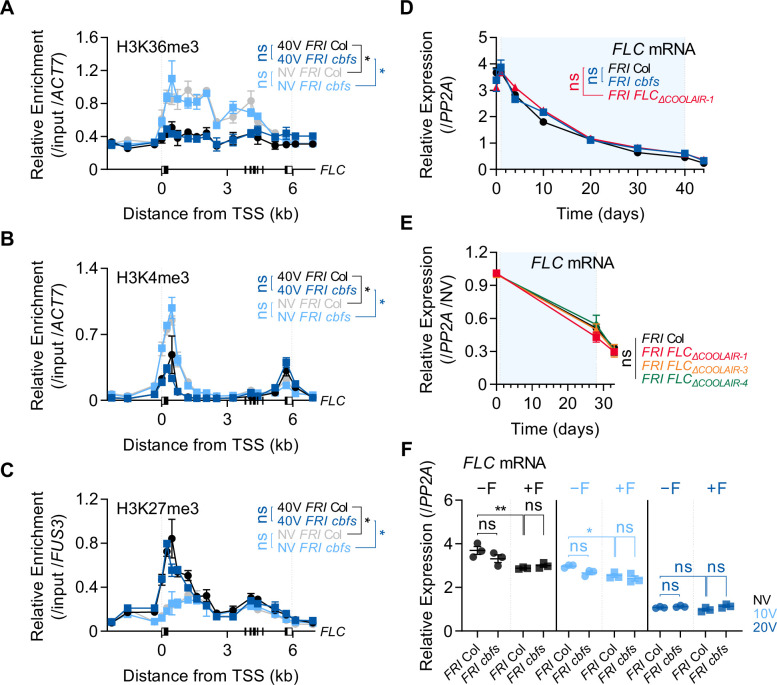
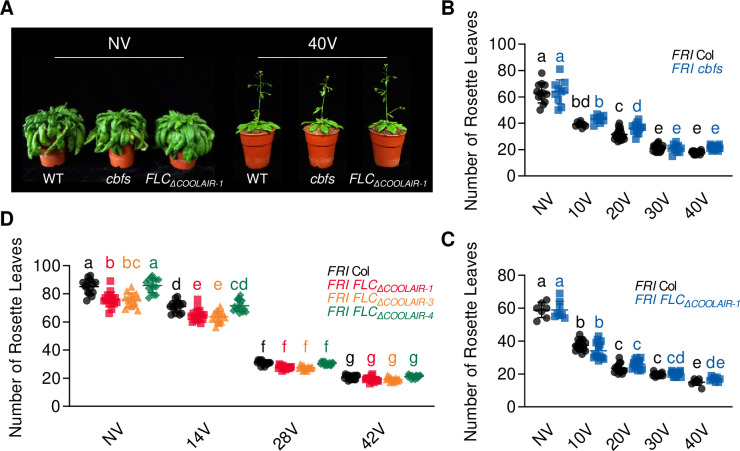
To synchronize flowering time with spring, many plants undergo vernalization, a floral-promotion process triggered by exposure to long-term winter cold. In , this is achieved through cold-mediated epigenetic silencing of the floral repressor, (). , a cold-induced antisense RNA transcribed from the locus, has been proposed to facilitate silencing. Here, we show that C-repeat (CRT)/dehydration-responsive elements (DREs) at the 3'-end of and CRT/DRE-binding factors (CBFs) are required for cold-mediated expression of . CBFs bind to CRT/DREs at the 3'-end of , both in vitro and in vivo, and CBF levels increase gradually during vernalization. Cold-induced expression is severely impaired in mutants in which all genes are knocked-out. Conversely, -overexpressing plants show increased levels even at warm temperatures. We show that is induced by CBFs during early stages of vernalization but levels decrease in later phases as chromatin transitions to an inactive state to which CBFs can no longer bind. We also demonstrate that and mutants exhibit a normal vernalization response despite their inability to activate expression during cold, revealing that is not required for the vernalization process.
为了使花期与春天同步,许多植物经历春化作用,这是一个由长期冬季寒冷触发的促进开花的过程。在拟南芥中,这是通过低温介导的花抑制因子( )的表观遗传沉默来实现的。有人提出,低温诱导的反义 RNA 从 基因转录,有助于 的沉默。在这里,我们表明, 的 3'末端的 CRT/脱水响应元件(DREs)和 CRT/DRE 结合因子(CBFs)对于低温介导的 的表达是必需的。CBFs 在体外和体内都与 3'末端的 CRT/DREs 结合,并且在春化过程中 CBF 水平逐渐增加。在所有 基因敲除的 突变体中,低温诱导的 表达严重受损。相反,过表达 的植物即使在温暖的温度下也表现出增加的 水平。我们表明,在春化的早期阶段,CBFs 诱导了 的表达,但随着 染色质向 CBFs 不再结合的非活性状态转变, 的水平在后期阶段下降。我们还证明,尽管在寒冷条件下不能激活 的表达, 和 突变体仍表现出正常的春化反应,这表明 对于春化过程不是必需的。