Computational and Systems Biology, John Innes Centre, Norwich Research Park, Norwich, United Kingdom.
Cell and Developmental Biology, John Innes Centre, Norwich Research Park, Norwich, United Kingdom.
Elife. 2023 Feb 27;12:e79408. doi: 10.7554/eLife.79408.
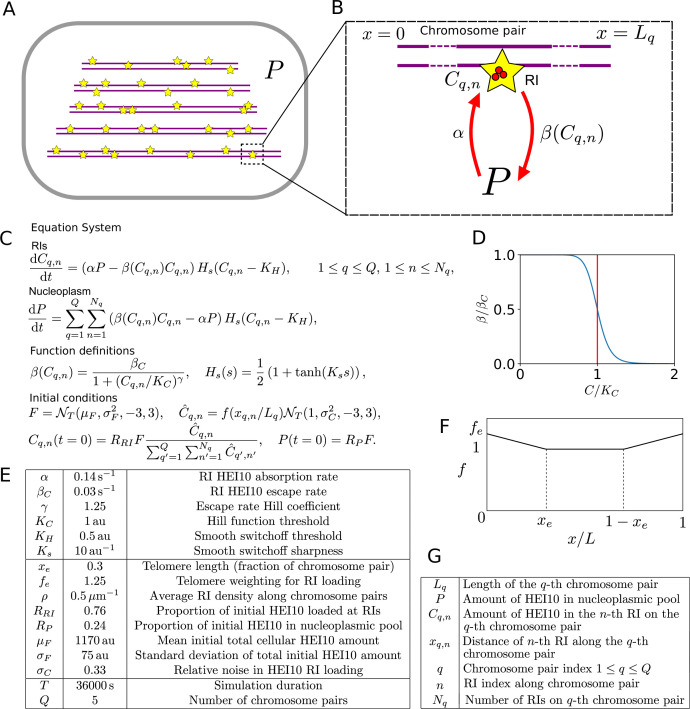
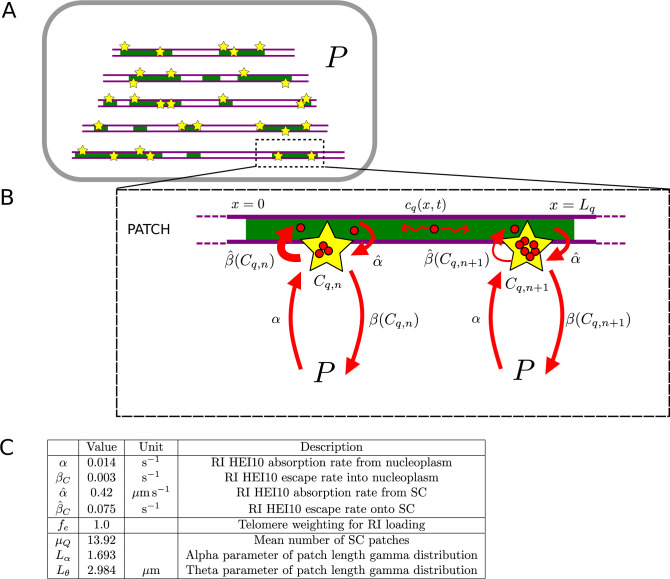
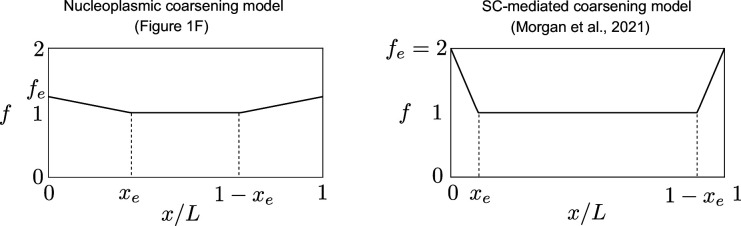
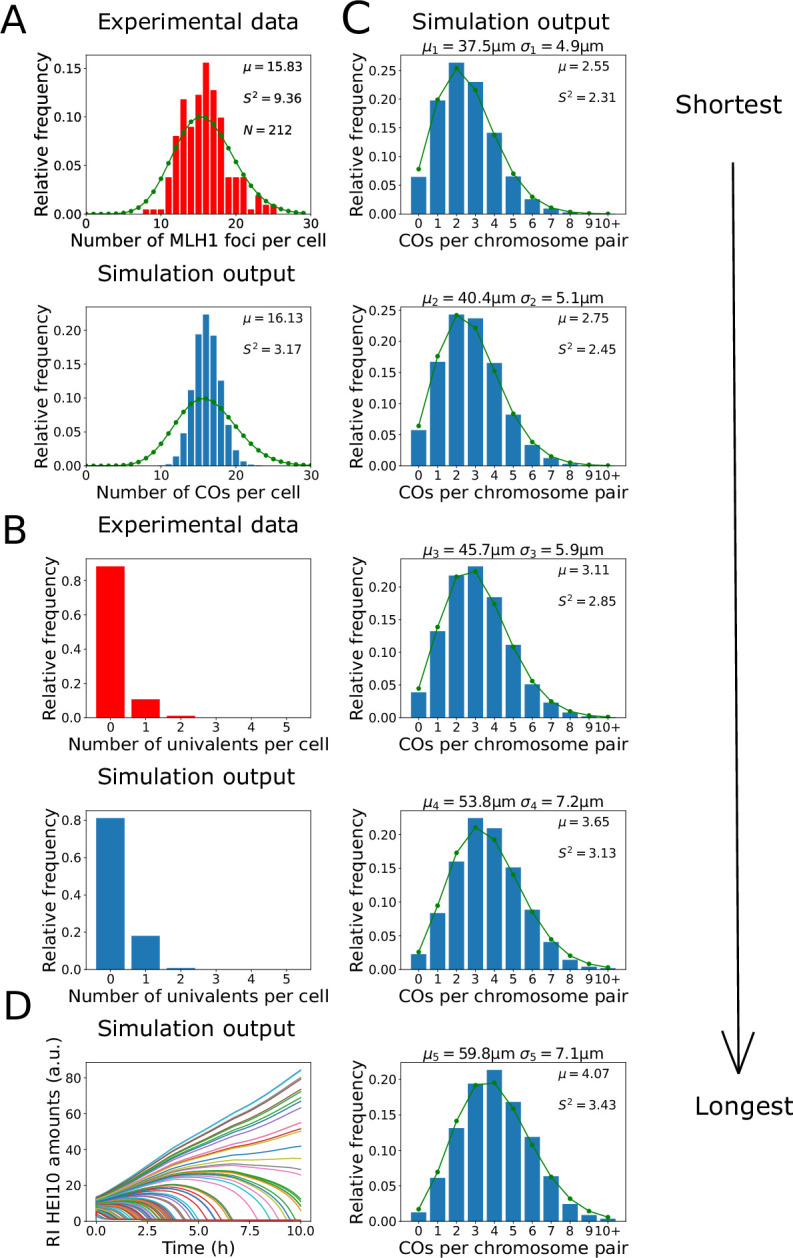
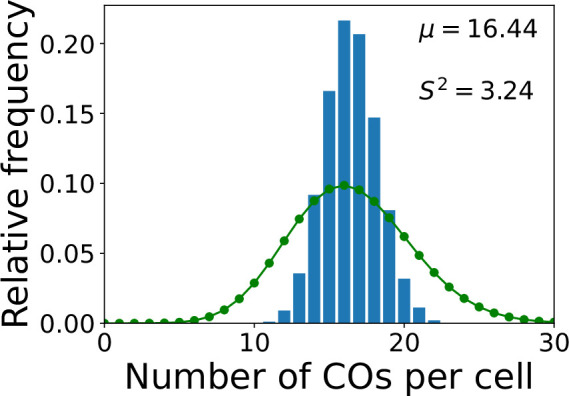
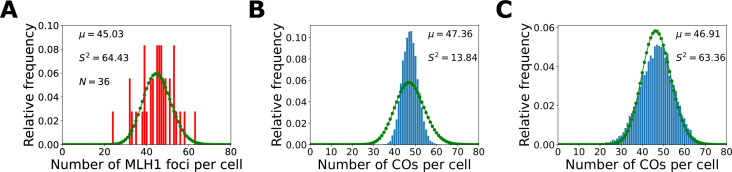
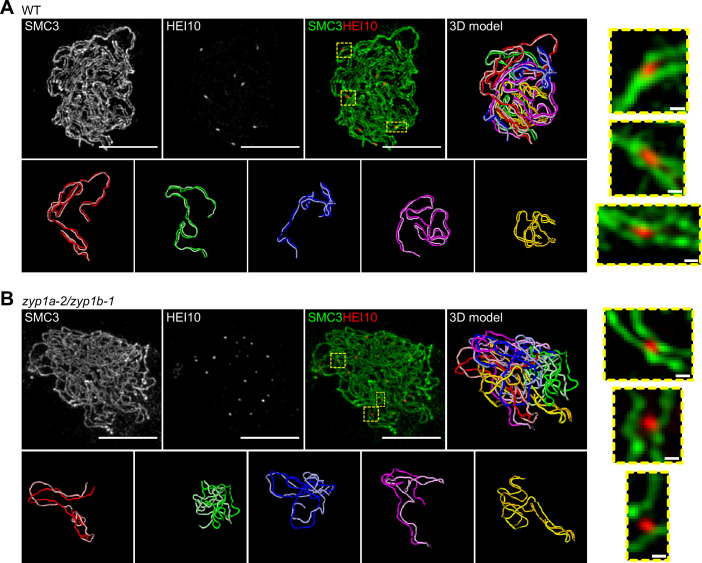
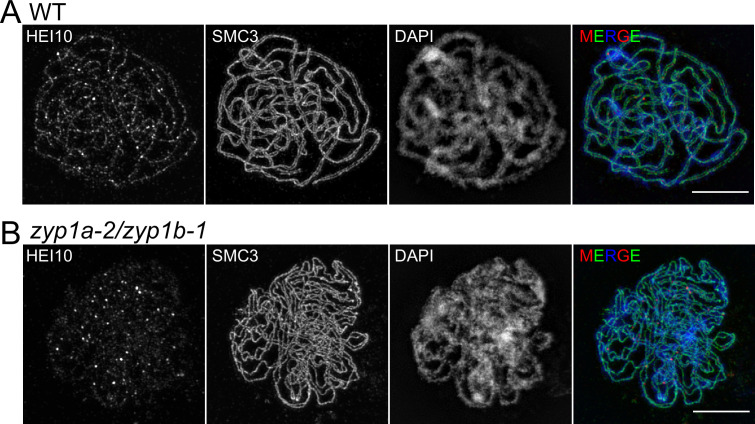
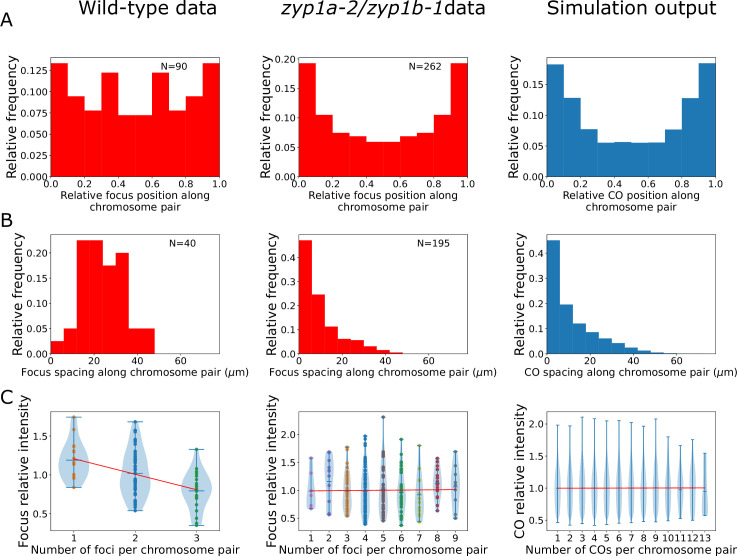
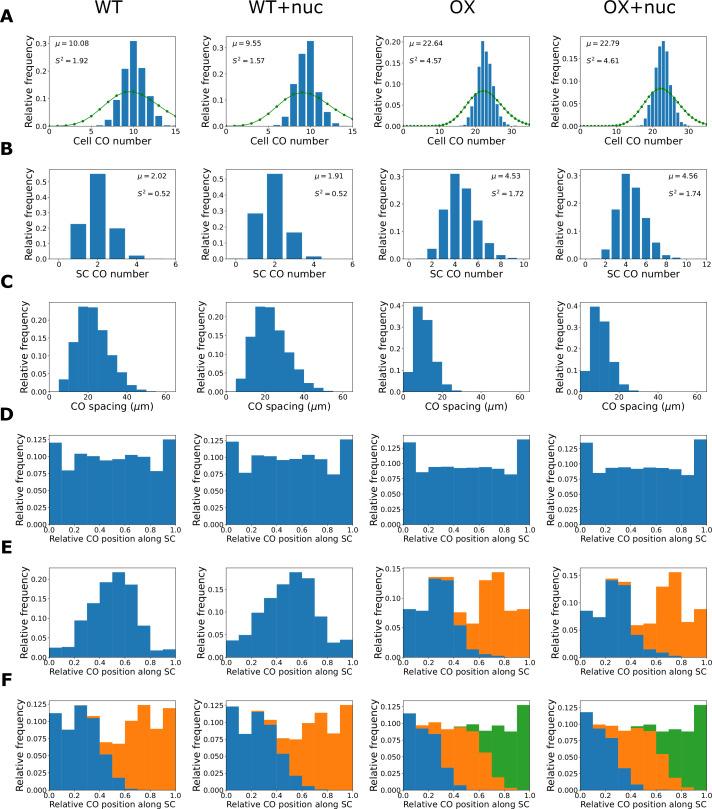
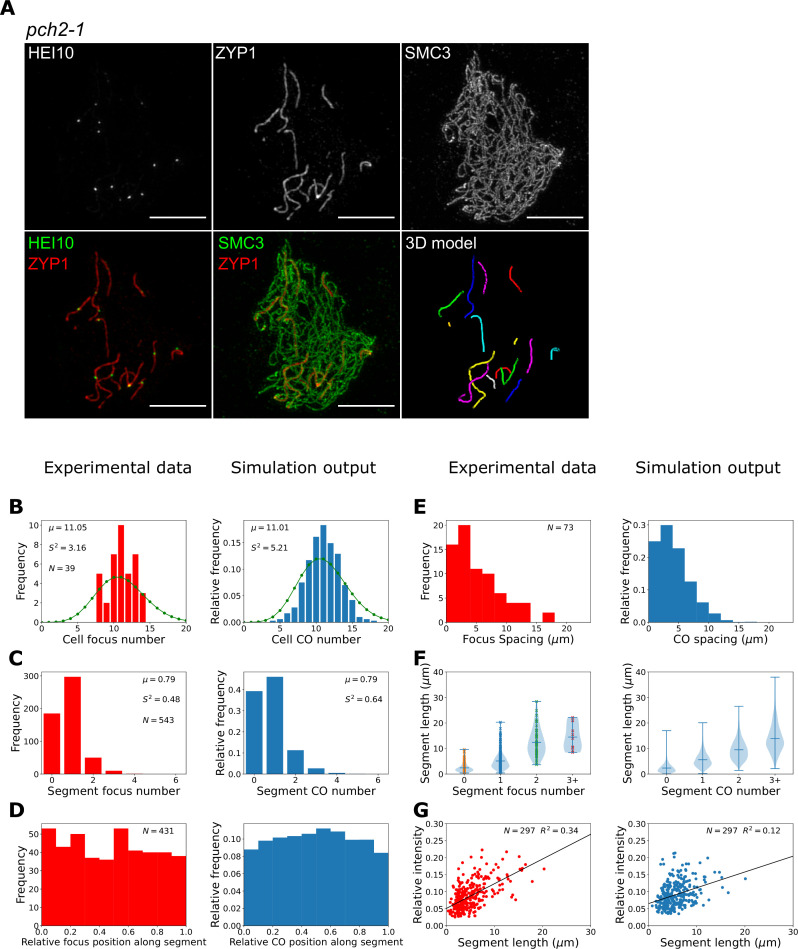
The shuffling of genetic material facilitated by meiotic crossovers is a critical driver of genetic variation. Therefore, the number and positions of crossover events must be carefully controlled. In an obligate crossover and repression of nearby crossovers on each chromosome pair are abolished in mutants that lack the synaptonemal complex (SC), a conserved protein scaffold. We use mathematical modelling and quantitative super-resolution microscopy to explore and mechanistically explain meiotic crossover pattering in lines with full, incomplete, or abolished synapsis. For mutants, which lack an SC, we develop a coarsening model in which crossover precursors globally compete for a limited pool of the pro-crossover factor HEI10, with dynamic HEI10 exchange mediated through the nucleoplasm. We demonstrate that this model is capable of quantitatively reproducing and predicting experimental crossover patterning and HEI10 foci intensity data. Additionally, we find that a model combining both SC- and nucleoplasm-mediated coarsening can explain crossover patterning in wild-type and in mutants, which display partial synapsis. Together, our results reveal that regulation of crossover patterning in wild-type and SC-defective mutants likely acts through the same underlying coarsening mechanism, differing only in the spatial compartments through which the pro-crossover factor diffuses.
减数分裂交叉重组所促进的遗传物质重排是遗传变异的关键驱动因素。因此,必须仔细控制交叉事件的数量和位置。在一个必需的交叉和抑制附近的交叉的情况下,在缺乏联会复合体(SC)的突变体中,每个染色体对的交叉事件都会被废除,SC 是一种保守的蛋白质支架。我们使用数学建模和定量超分辨率显微镜来探索和从机制上解释在具有完整、不完整或废除联会的系中减数分裂交叉模式。对于缺乏 SC 的突变体,我们开发了一个粗化模型,其中交叉前体在有限的前交叉因子 HEI10 池中全局竞争,通过核质介导动态 HEI10 交换。我们证明,该模型能够定量重现和预测实验交叉模式和 HEI10 焦点强度数据。此外,我们发现,一个结合了 SC 和核质介导的粗化的模型可以解释野生型和显示部分联会的突变体中的交叉模式。总之,我们的结果表明,野生型和 SC 缺陷突变体中交叉模式的调节可能通过相同的基础粗化机制起作用,只是前交叉因子扩散的空间隔室不同。