Department of Microbiology, Immunology and Molecular Genetics, University of Kentucky College of Medicine, Lexington, KY, United States.
Department of Pharmacology and Nutritional Science, University of Kentucky College of Medicine, Lexington, KY, United States.
Front Cell Infect Microbiol. 2023 Feb 15;13:1110508. doi: 10.3389/fcimb.2023.1110508. eCollection 2023.
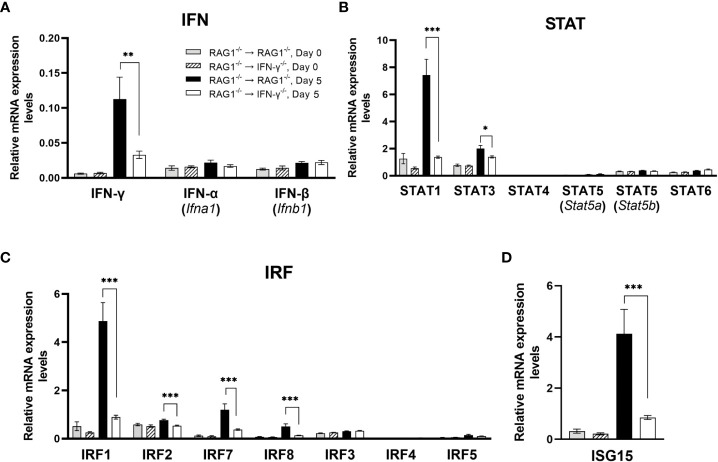
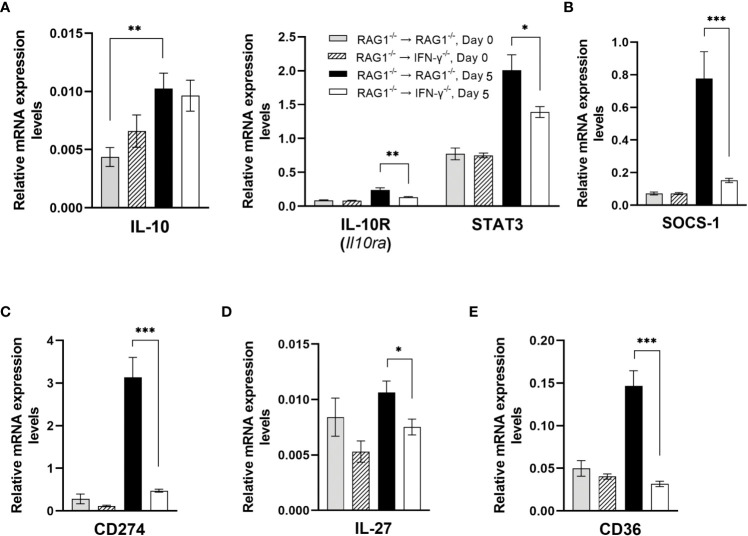
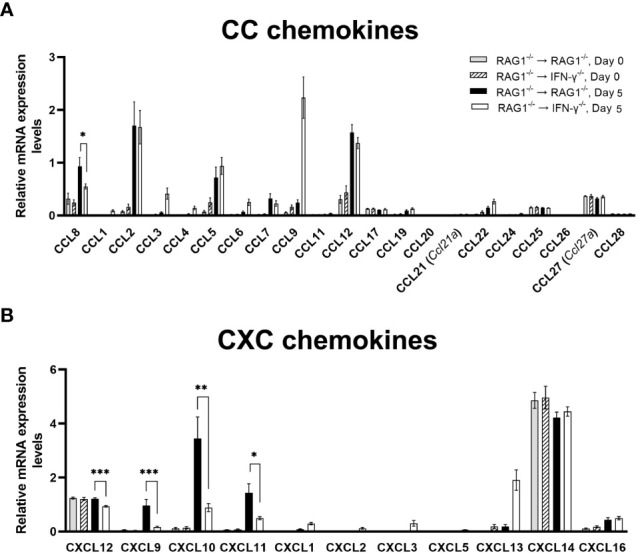
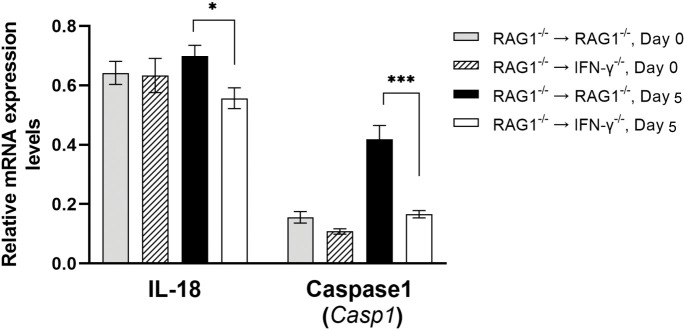
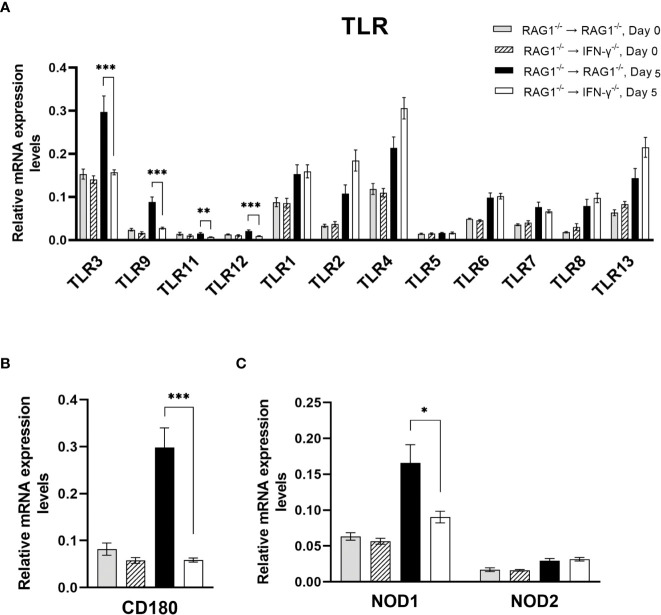
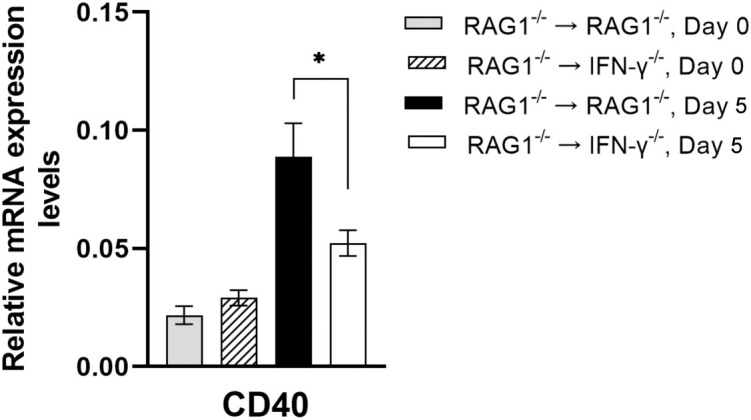
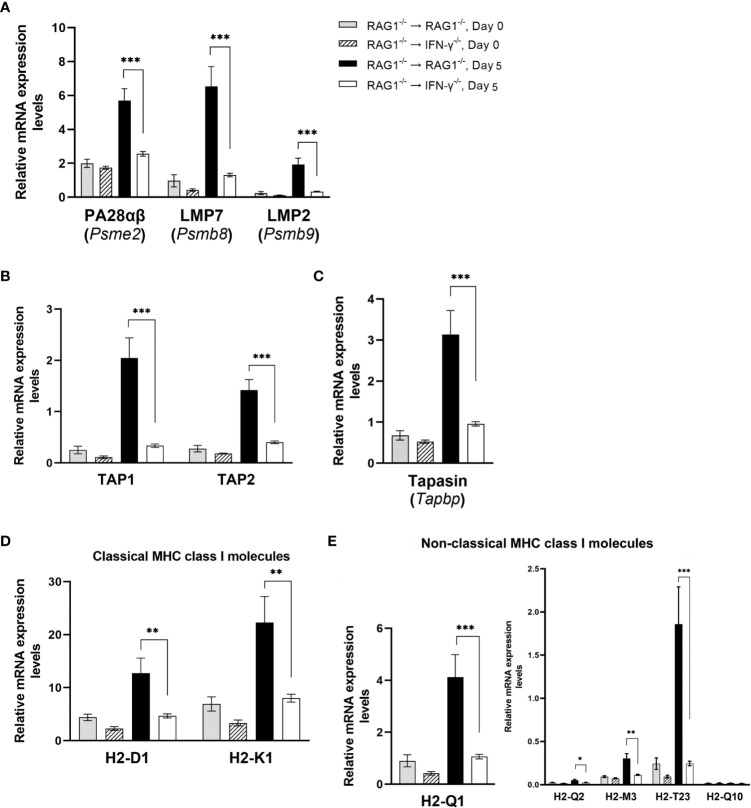
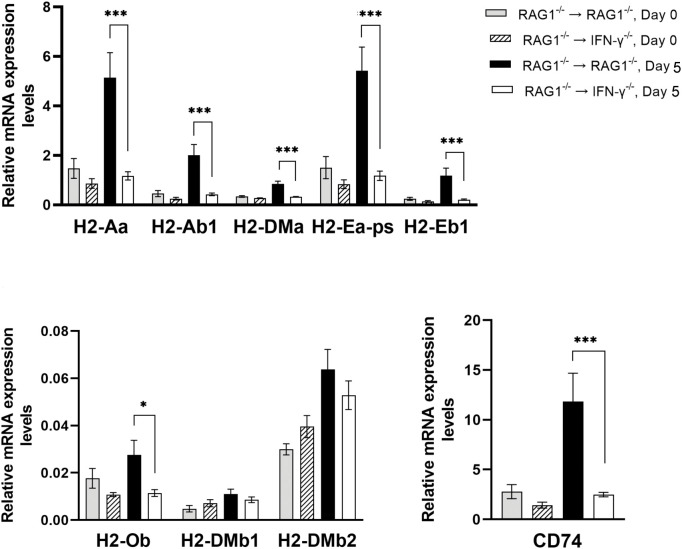
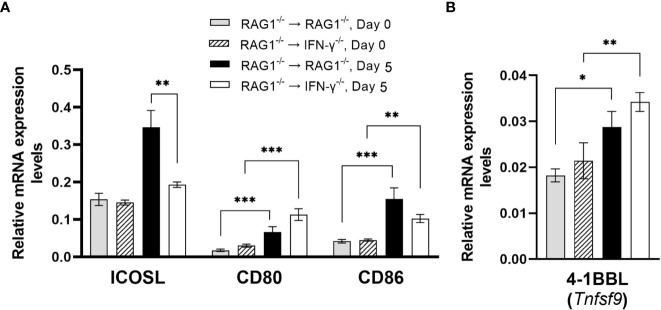
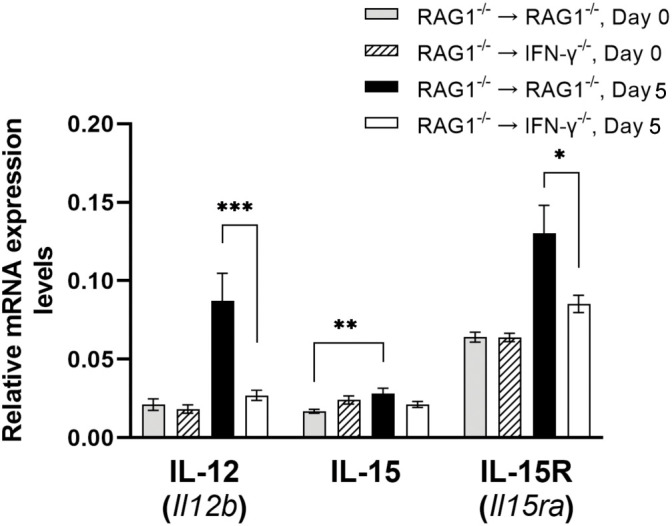
We previously demonstrated that brain-resident cells produce IFN-γ in response to reactivation of cerebral infection with . To obtain an overall landscape view of the effects of IFN-γ from brain-resident cells on the cerebral protective immunity, in the present study we employed NanoString nCounter assay and quantified mRNA levels for 734 genes in myeloid immunity in the brains of T and B cell-deficient, bone marrow chimeric mice with and without IFN-γ production by brain-resident cells in response to reactivation of cerebral infection. Our study revealed that IFN-γ produced by brain-resident cells amplified mRNA expression for the molecules to activate the protective innate immunity including 1) chemokines for recruitment of microglia and macrophages (CCL8 and CXCL12) and 2) the molecules for activating those phagocytes (IL-18, TLRs, NOD1, and CD40) for killing tachyzoites. Importantly, IFN-γ produced by brain-resident cells also upregulated cerebral expression of molecules for facilitating the protective T cell immunity, which include the molecules for 1) recruiting effector T cells (CXCL9, CXCL10, and CXCL11), 2) antigen processing (PA28αβ, LMP2, and LMP7), transporting the processed peptides (TAP1 and TAP2), assembling the transported peptides to the MHC class I molecules (Tapasin), and the MHC class I (H2-K1 and H2-D1) and Ib molecules (H2-Q1, H-2Q2, and H2-M3) for presenting antigens to activate the recruited CD8 T cells, 3) MHC class II molecules (H2-Aa, H2-Ab1, H2-Eb1, H2-Ea-ps, H2-DMa, H2-Ob, and CD74) to present antigens for CD4 T cell activation, 4) co-stimulatory molecules (ICOSL) for T cell activation, and 5) cytokines (IL-12, IL-15, and IL-18) facilitating IFN-γ production by NK and T cells. Notably, the present study also revealed that IFN-γ production by brain-resident cells also upregulates cerebral expressions of mRNA for the downregulatory molecules (IL-10, STAT3, SOCS1, CD274 [PD-L1], IL-27, and CD36), which can prevent overly stimulated IFN-γ-mediated pro-inflammatory responses and tissue damages. Thus, the present study uncovered the previously unrecognized the capability of IFN-γ production by brain-resident cells to upregulate expressions of a wide spectrum of molecules for coordinating both innate and T cell-mediated protective immunity with a fine-tuning regulation system to effectively control cerebral infection with .
我们之前证明,大脑驻留细胞在大脑感染重新激活时会产生 IFN-γ。为了全面了解大脑驻留细胞产生的 IFN-γ对大脑保护性免疫的影响,在本研究中,我们采用 NanoString nCounter 检测方法,定量分析了 T 和 B 细胞缺陷、骨髓嵌合小鼠大脑中髓系免疫中 734 种基因的 mRNA 水平,这些小鼠在大脑驻留细胞重新激活大脑感染时产生 IFN-γ。我们的研究表明,大脑驻留细胞产生的 IFN-γ放大了激活保护性先天免疫的分子的 mRNA 表达,包括:1)招募小胶质细胞和巨噬细胞的趋化因子(CCL8 和 CXCL12),2)激活这些吞噬细胞的分子(IL-18、TLRs、NOD1 和 CD40)以杀死速殖子。重要的是,大脑驻留细胞产生的 IFN-γ也上调了大脑中促进保护性 T 细胞免疫的分子表达,包括:1)募集效应 T 细胞的分子(CXCL9、CXCL10 和 CXCL11),2)抗原加工分子(PA28αβ、LMP2 和 LMP7),转运加工肽的分子(TAP1 和 TAP2),将转运肽组装到 MHC I 分子(Tapasin)上的分子,以及 MHC I(H2-K1 和 H2-D1)和 Ib 分子(H2-Q1、H-2Q2 和 H2-M3)以将抗原呈递给激活募集的 CD8 T 细胞,3)MHC II 分子(H2-Aa、H2-Ab1、H2-Eb1、H2-Ea-ps、H2-DMa、H2-Ob 和 CD74)以将抗原呈递给 CD4 T 细胞激活,4)共刺激分子(ICOSL)用于 T 细胞激活,5)细胞因子(IL-12、IL-15 和 IL-18)促进 NK 和 T 细胞产生 IFN-γ。值得注意的是,本研究还揭示了大脑驻留细胞产生的 IFN-γ还上调了大脑中下调分子(IL-10、STAT3、SOCS1、CD274 [PD-L1]、IL-27 和 CD36)的 mRNA 表达,这可以防止过度刺激 IFN-γ 介导的促炎反应和组织损伤。因此,本研究揭示了大脑驻留细胞产生 IFN-γ的先前未被认识的能力,即上调一系列分子的表达,以协调先天免疫和 T 细胞介导的保护性免疫,并通过精细调节系统有效控制大脑感染。