Department of Psychiatry and Psychotherapy, Medical Center - University of Freiburg, Faculty of Medicine, University of Freiburg, Freiburg, Germany.
Department of Physiology, Anatomy and Genetics, University of Oxford, Oxford, United Kingdom.
Elife. 2023 Mar 9;12:e84740. doi: 10.7554/eLife.84740.
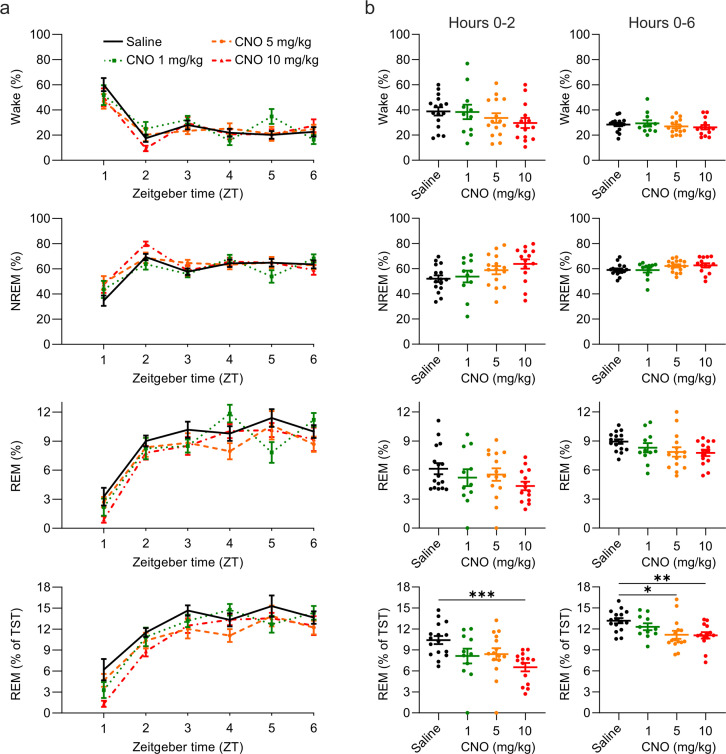
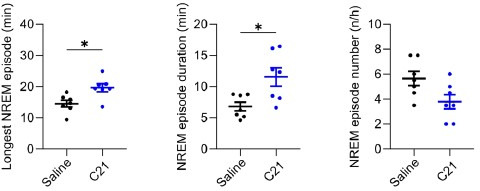
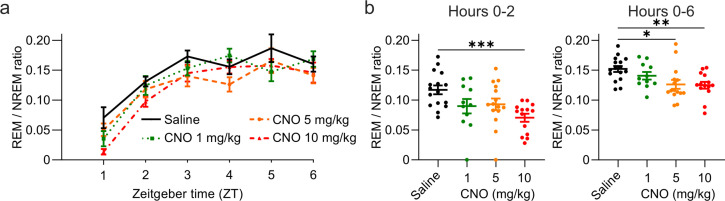
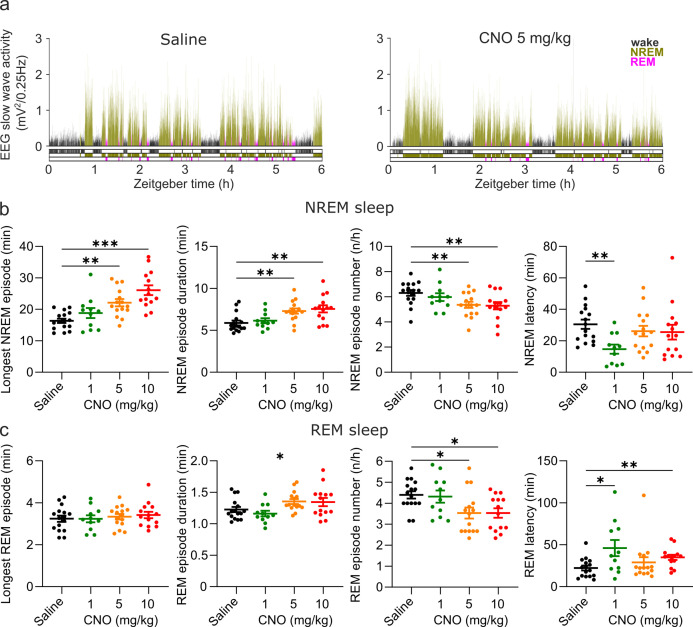
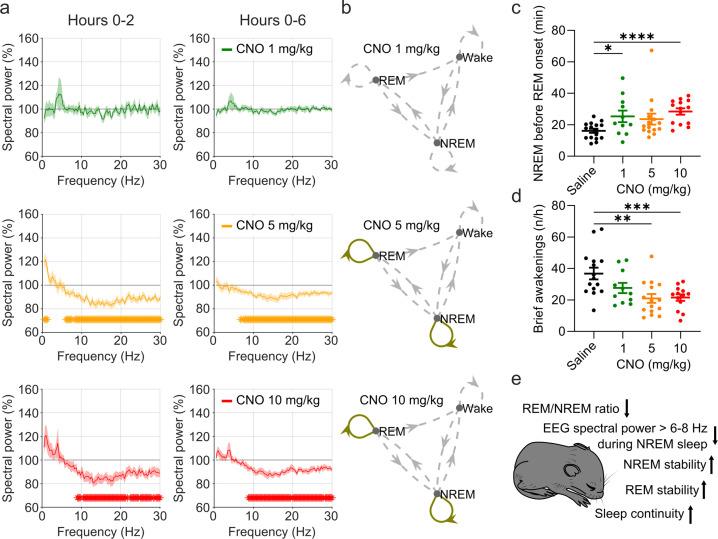
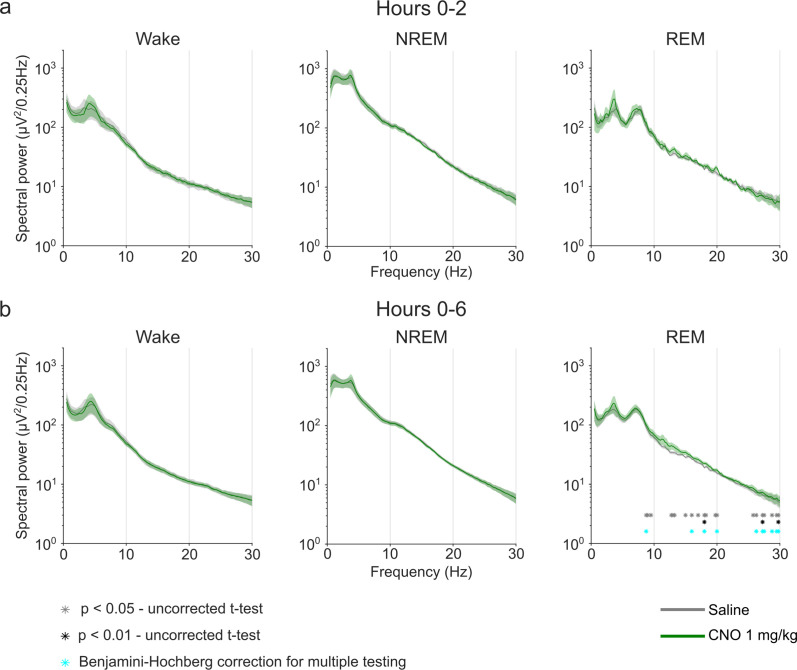
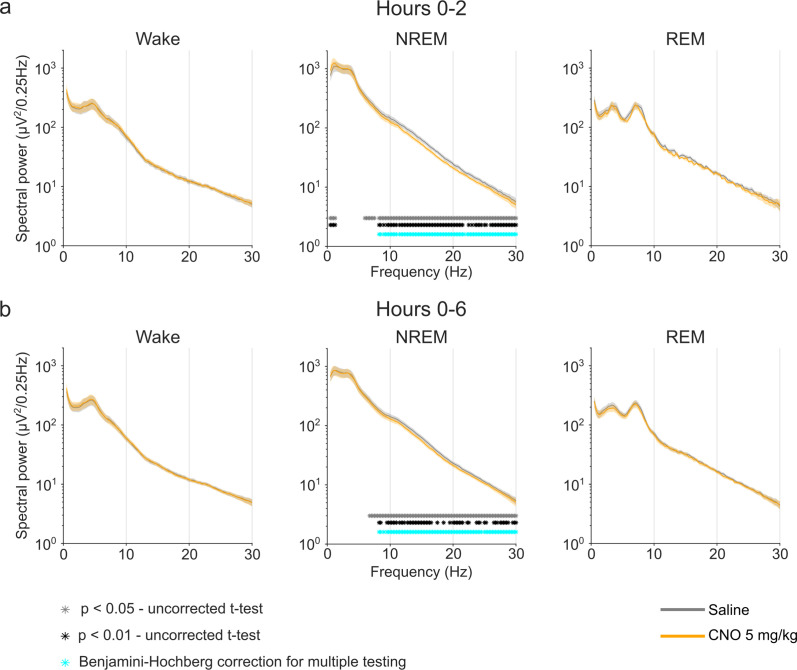
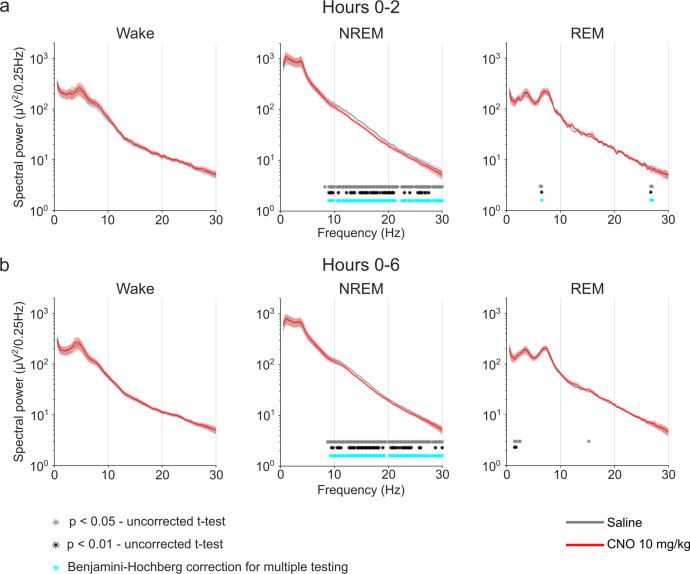
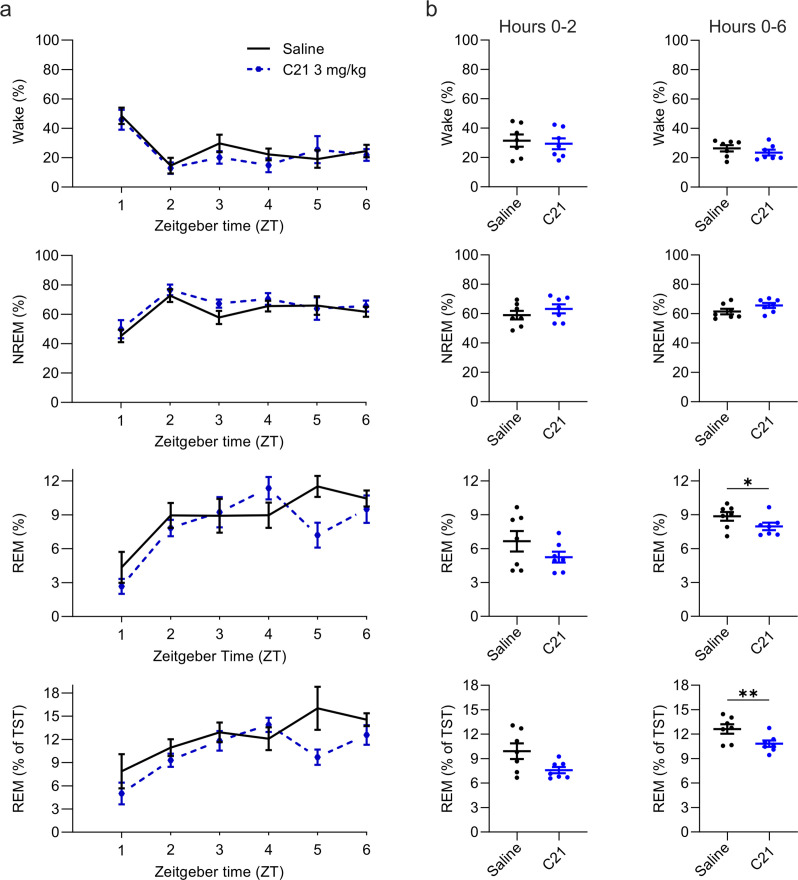
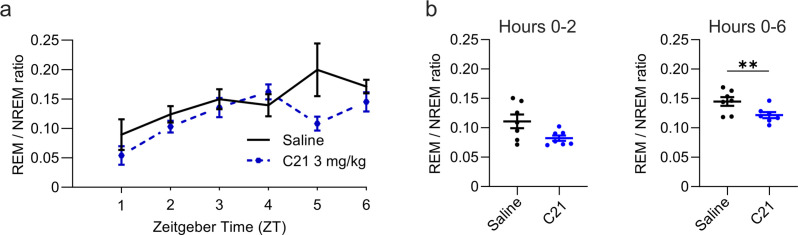
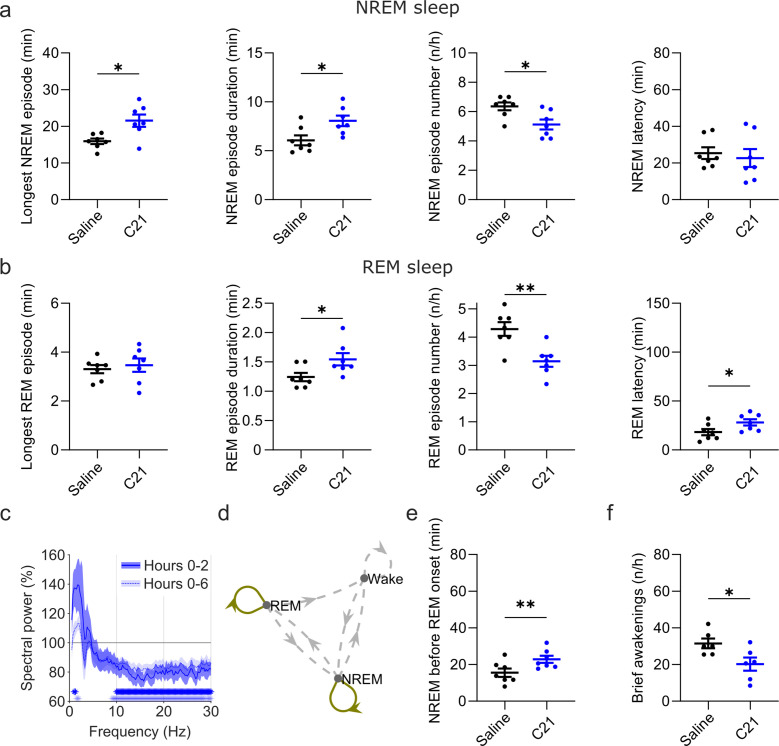
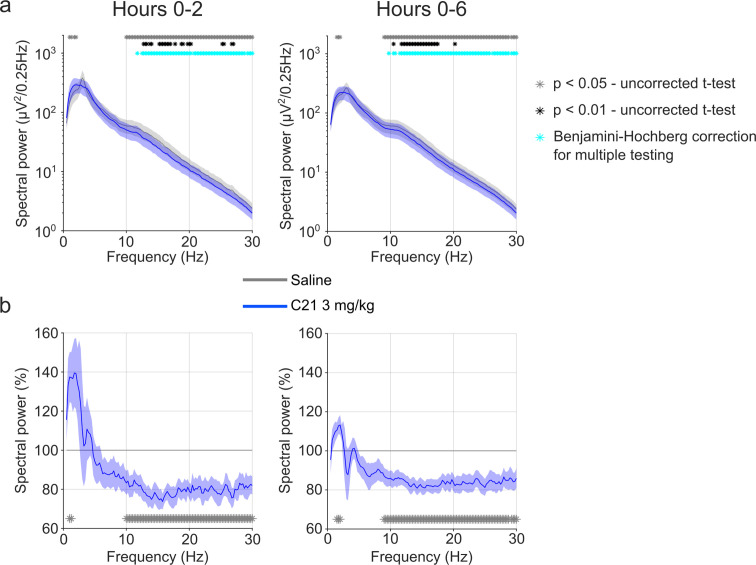
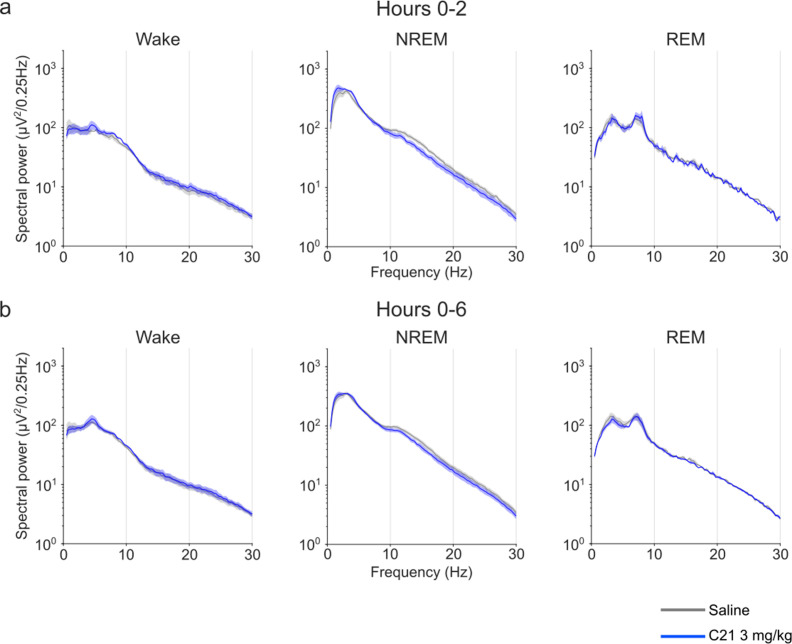
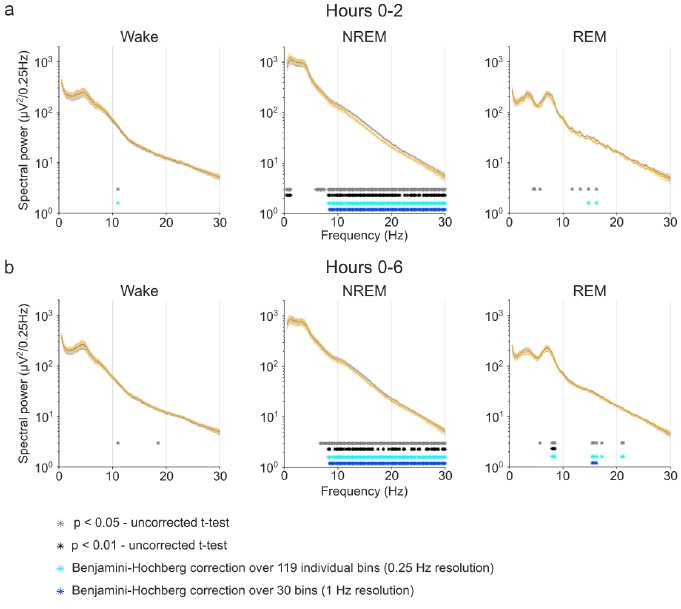
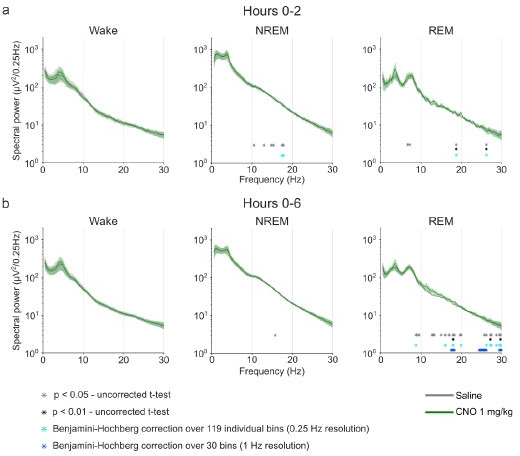
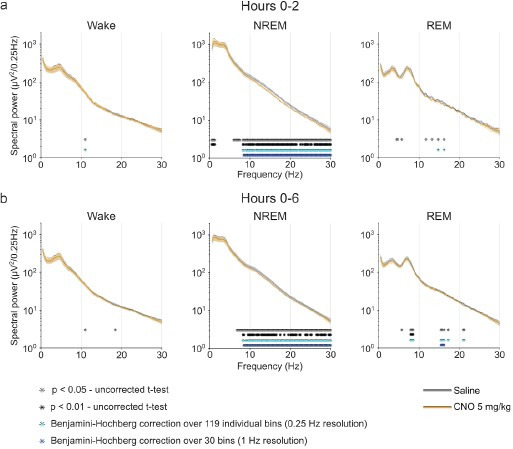
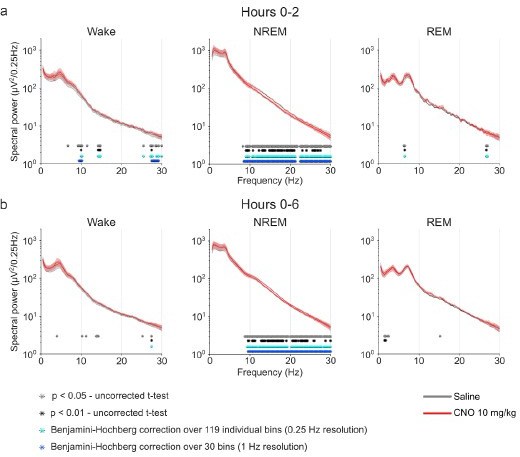
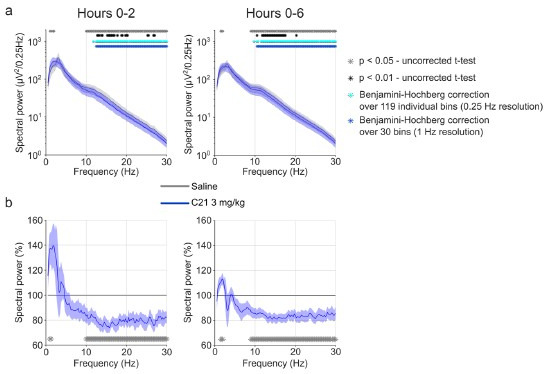
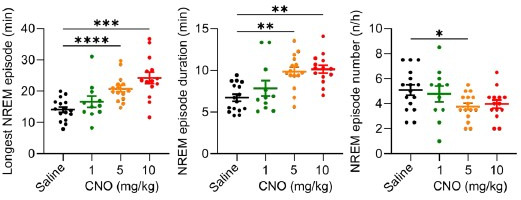
Designer receptors exclusively activated by designer drugs (DREADDs) are chemogenetic tools for remote control of targeted cell populations using chemical actuators that bind to modified receptors. Despite the popularity of DREADDs in neuroscience and sleep research, potential effects of the DREADD actuator clozapine-N-oxide (CNO) on sleep have never been systematically tested. Here, we show that intraperitoneal injections of commonly used CNO doses (1, 5, and 10 mg/kg) alter sleep in wild-type male laboratory mice. Using electroencephalography (EEG) and electromyography (EMG) to analyse sleep, we found a dose-dependent suppression of rapid eye movement (REM) sleep, changes in EEG spectral power during non-REM (NREM) sleep, and altered sleep architecture in a pattern previously reported for clozapine. Effects of CNO on sleep could arise from back-metabolism to clozapine or binding to endogenous neurotransmitter receptors. Interestingly, we found that the novel DREADD actuator, compound 21 (C21, 3 mg/kg), similarly modulates sleep despite a lack of back-metabolism to clozapine. Our results demonstrate that both CNO and C21 can modulate sleep of mice not expressing DREADD receptors. This implies that back-metabolism to clozapine is not the sole mechanism underlying side effects of chemogenetic actuators. Therefore, any chemogenetic experiment should include a DREADD-free control group injected with the same CNO, C21, or newly developed actuator. We suggest that electrophysiological sleep assessment could serve as a sensitive tool to test the biological inertness of novel chemogenetic actuators.
设计者受体专门由设计者药物激活(DREADDs)是一种化学遗传工具,用于使用化学激活剂远程控制靶向细胞群体,化学激活剂与修饰后的受体结合。尽管 DREADDs 在神经科学和睡眠研究中很受欢迎,但 DREADD 激活剂氯氮平-N-氧化物(CNO)对睡眠的潜在影响从未被系统测试过。在这里,我们表明,腹腔内注射常用的 CNO 剂量(1、5 和 10mg/kg)会改变野生型雄性实验室小鼠的睡眠。使用脑电图(EEG)和肌电图(EMG)分析睡眠,我们发现快速眼动(REM)睡眠的剂量依赖性抑制,非快速眼动(NREM)睡眠期间 EEG 频谱功率的变化,以及睡眠结构的改变,这些变化模式与氯氮平以前的报道一致。CNO 对睡眠的影响可能来自于氯氮平的反向代谢产物或与内源性神经递质受体的结合。有趣的是,我们发现新型 DREADD 激活剂化合物 21(C21,3mg/kg)也能调节睡眠,尽管没有氯氮平的反向代谢产物。我们的结果表明,CNO 和 C21 都可以调节不表达 DREADD 受体的小鼠的睡眠。这意味着氯氮平的反向代谢产物不是化学遗传激活剂副作用的唯一机制。因此,任何化学遗传实验都应该包括一个没有 DREADD 的对照组,用相同的 CNO、C21 或新开发的激活剂注射。我们建议电生理睡眠评估可以作为测试新型化学遗传激活剂生物惰性的敏感工具。