Gill Kevin S, Mehta Kritika, Heredia Jeremiah D, Krishnamurthy Vishnu V, Zhang Kai, Procko Erik
Department of Biochemistry, University of Illinois, Urbana, IL 61801, USA.
Current affiliation: Codexis, Redwood City, CA 94063.
bioRxiv. 2023 Mar 25:2023.03.25.534231. doi: 10.1101/2023.03.25.534231.
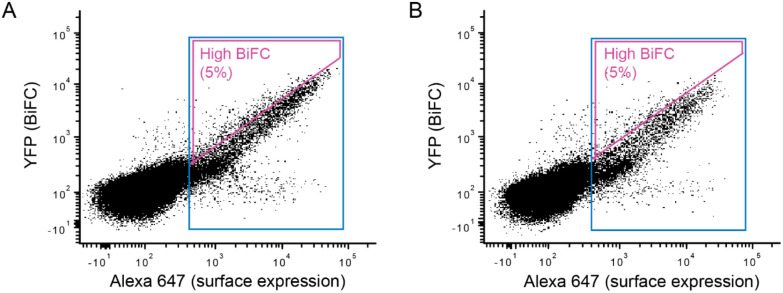
Chemokine receptors are members of the rhodopsin-like class A GPCRs whose signaling through G proteins drives the directional movement of cells in response to a chemokine gradient. Chemokine receptors CXCR4 and CCR5 have been extensively studied due to their roles in white blood cell development and inflammation and their status as coreceptors for HIV-1 infection, among other functions. Both receptors form dimers or oligomers but the function/s of self-associations are unclear. While CXCR4 has been crystallized in a dimeric arrangement, available atomic resolution structures of CCR5 are monomeric. To investigate the dimerization interfaces of these chemokine receptors, we used a bimolecular fluorescence complementation (BiFC)-based screen and deep mutational scanning to find mutations that modify receptor self-association. Many disruptive mutations promoted self-associations nonspecifically, suggesting they aggregated in the membrane. A mutationally intolerant region was found on CXCR4 that matched the crystallographic dimer interface, supporting this dimeric arrangement in living cells. A mutationally intolerant region was also observed on the surface of CCR5 by transmembrane helices 3 and 4. Mutations from the deep mutational scan that reduce BiFC were validated and were localized in the transmembrane domains as well as the C-terminal cytoplasmic tails where they reduced lipid microdomain localization. The reduced self-association mutants of CXCR4 had increased binding to the ligand CXCL12 but diminished calcium signaling. There was no change in syncytia formation with cells expressing HIV-1 Env. The data highlight that multiple mechanisms are involved in self-association of chemokine receptor chains.
趋化因子受体是视紫红质样A类G蛋白偶联受体(GPCR)的成员,其通过G蛋白进行信号传导,驱动细胞响应趋化因子梯度进行定向移动。趋化因子受体CXCR4和CCR5因其在白细胞发育和炎症中的作用以及作为HIV-1感染共受体的地位等功能而受到广泛研究。两种受体均形成二聚体或寡聚体,但自缔合的功能尚不清楚。虽然CXCR4已以二聚体形式结晶,但CCR5的现有原子分辨率结构为单体。为了研究这些趋化因子受体的二聚化界面,我们使用基于双分子荧光互补(BiFC)的筛选和深度突变扫描来寻找改变受体自缔合的突变。许多破坏性突变非特异性地促进了自缔合,表明它们在膜中聚集。在CXCR4上发现了一个与晶体学二聚体界面匹配的突变耐受区域,支持其在活细胞中的这种二聚体排列。在CCR5跨膜螺旋3和4的表面也观察到一个突变耐受区域。深度突变扫描中降低BiFC的突变得到验证,并定位在跨膜结构域以及C端细胞质尾巴中,在这些位置它们减少了脂质微区定位。CXCR4的自缔合减少突变体与配体CXCL12的结合增加,但钙信号传导减弱。与表达HIV-1 Env的细胞形成合胞体的情况没有变化。数据突出表明趋化因子受体链的自缔合涉及多种机制。