Section of Cell Biology and Biophysics, Department of Biology, School of Science, National and Kapodistrian University of Athens (NKUA), 15701 Athens, Greece.
Int J Mol Sci. 2023 Jun 2;24(11):9676. doi: 10.3390/ijms24119676.
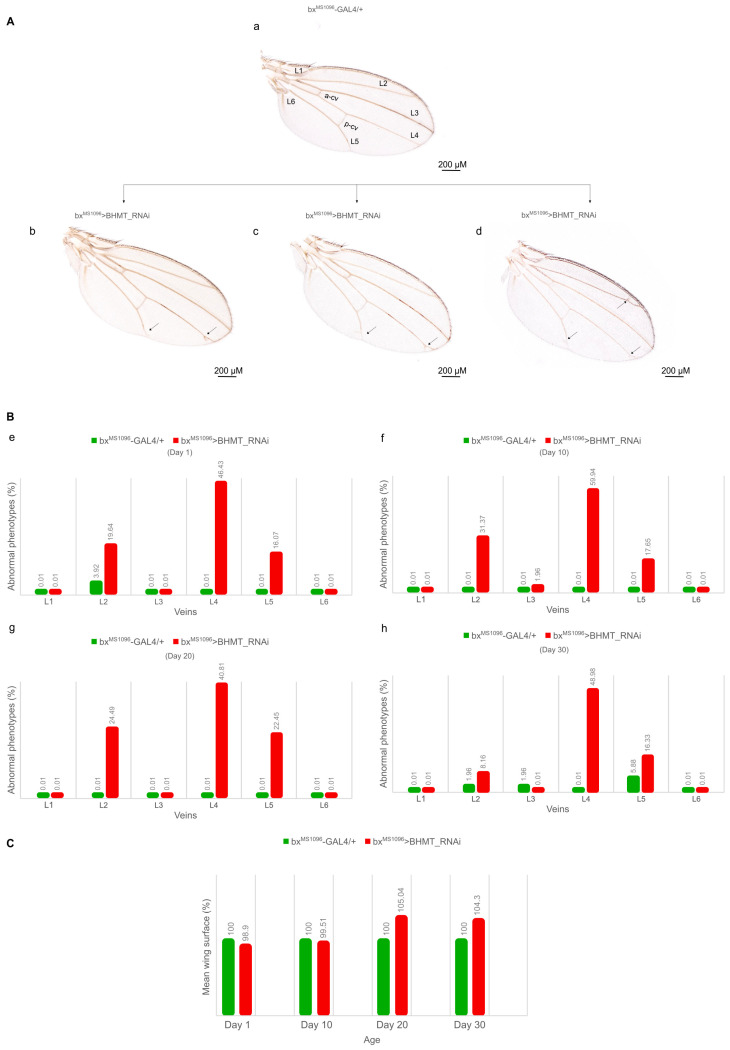
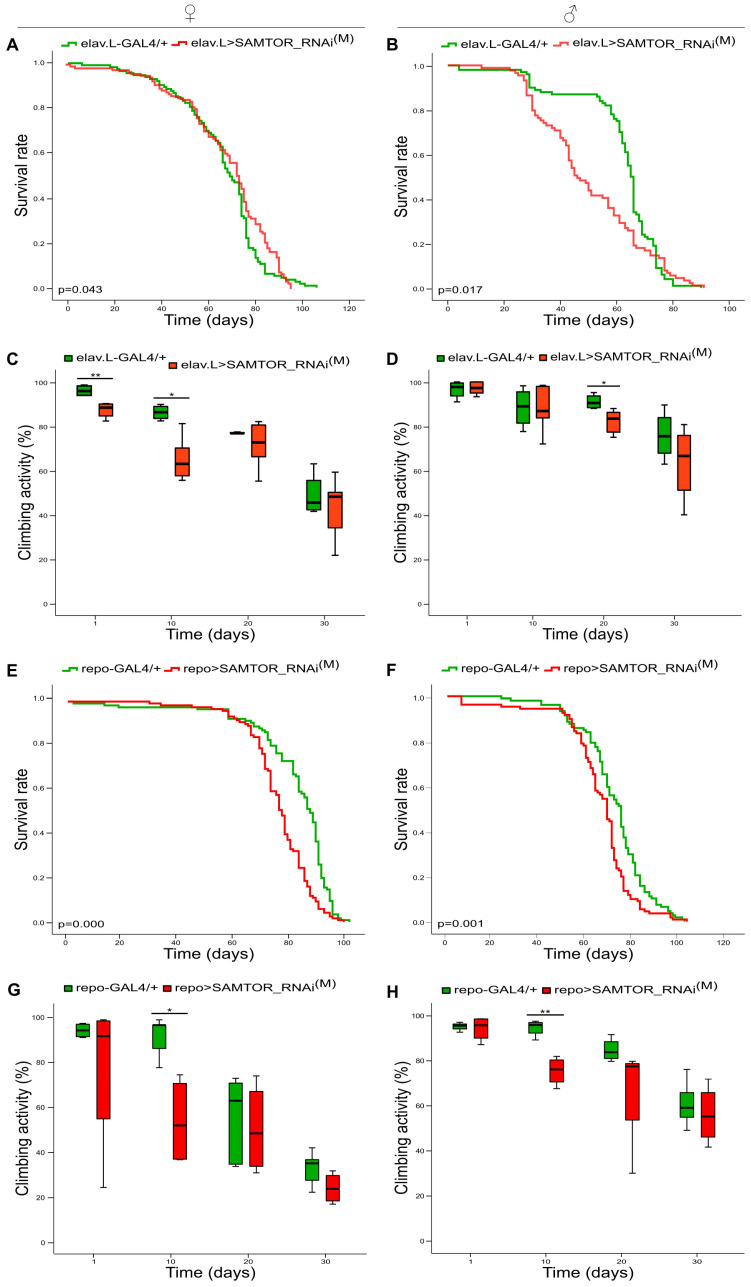
mTORC1 regulates mammalian cell metabolism and growth in response to diverse environmental stimuli. Nutrient signals control the localization of mTORC1 onto lysosome surface scaffolds that are critically implicated in its amino acid-dependent activation. Arginine, leucine and S-adenosyl-methionine (SAM) can serve as major mTORC1-signaling activators, with SAM binding to SAMTOR (SAM + TOR), a fundamental SAM sensor, preventing the protein's (SAMTOR's) inhibitory action(s) against mTORC1, thereby triggering its (mTORC1) kinase activity. Given the lack of knowledge regarding the role of SAMTOR in invertebrates, we have identified the SAMTOR homologue (dSAMTOR) in silico and have, herein, genetically targeted it through the utilization of the GAL4/UAS transgenic tool. Survival profiles and negative geotaxis patterns were examined in both control and -downregulated adult flies during aging. One of the two gene-targeted schemes resulted in lethal phenotypes, whereas the other one caused rather moderate pathologies in most tissues. The screening of head-specific kinase activities, via PamGene technology application, unveiled the significant upregulation of several kinases, including the dTORC1 characteristic substrate dp70S6K, in -downregulated flies, thus strongly supporting the inhibitory dSAMTOR action(s) upon the dTORC1/dp70S6K signaling axis in brain settings. Importantly, genetic targeting of the BHMT bioinformatics counterpart (dBHMT), an enzyme that catabolizes betaine to produce methionine (the SAM precursor), led to severe compromises in terms of fly longevity, with glia-, motor neuron- and muscle-specific dBHMT downregulations exhibiting the strongest effects. Abnormalities in wing vein architectures were also detected in -targeted flies, thereby justifying their notably reduced negative geotaxis capacities herein observed mainly in the brain-(mid)gut axis. In vivo adult fly exposure to clinically relevant doses of methionine revealed the mechanistic synergism of decreased dSAMTOR and increased methionine levels in pathogenic longevity, thus rendering (d)SAMTOR an important component in methionine-associated disorders, including homocystinuria(s).
mTORC1 响应各种环境刺激调节哺乳动物细胞代谢和生长。营养信号控制 mTORC1 定位于溶酶体表面支架上,这对于其氨基酸依赖性激活至关重要。精氨酸、亮氨酸和 S-腺苷甲硫氨酸 (SAM) 可以作为 mTORC1 信号激活的主要物质,SAM 结合到 SAMTOR(SAM + TOR),这是一种基本的 SAM 传感器,防止蛋白质(SAMTOR)对 mTORC1 的抑制作用,从而触发其(mTORC1)激酶活性。鉴于缺乏关于 SAMTOR 在无脊椎动物中的作用的知识,我们通过使用 GAL4/UAS 转基因工具在计算机上鉴定了 SAMTOR 同源物(dSAMTOR),并在此对其进行了基因靶向。在衰老过程中,通过检查对照和下调成年果蝇的生存曲线和负趋地性模式来研究。两种基因靶向方案中的一种导致致命表型,而另一种在大多数组织中导致相当温和的病变。通过 PamGene 技术应用筛选头特异性激酶活性,揭示了几种激酶的显著上调,包括 dTORC1 特征底物 dp70S6K,在下调的果蝇中,这强烈支持在大脑环境中 dSAMTOR 对 dTORC1/dp70S6K 信号轴的抑制作用。重要的是,BHMT 生物信息学对应物(dBHMT)的基因靶向,该酶将甜菜碱分解为产生蛋氨酸(SAM 前体)的酶,导致果蝇寿命严重受损,神经胶质、运动神经元和肌肉特异性 dBHMT 下调表现出最强的影响。在下调的果蝇中还检测到翅膀静脉结构的异常,从而证明了它们在此处观察到的负趋地性能力的明显降低,主要在大脑-(中)肠轴中。体内成年果蝇暴露于临床相关剂量的蛋氨酸揭示了降低的 dSAMTOR 和增加的蛋氨酸水平在发病寿命中的机制协同作用,从而使(d)SAMTOR 成为与蛋氨酸相关疾病(包括高胱氨酸尿症)的重要组成部分。