Kinases, Protein Phosphorylation and Cancer Group, Structural Biology Programme, Spanish National Cancer Research Center (CNIO), C/Melchor Fernández Almagro num. 3, 28029, Madrid, Spain.
Faculty of Experimental Sciences, Universidad Francisco de Vitoria (UFV), 28223, Pozuelo de Alarcón, Madrid, Spain.
Nat Commun. 2023 Oct 17;14(1):6548. doi: 10.1038/s41467-023-41890-7.
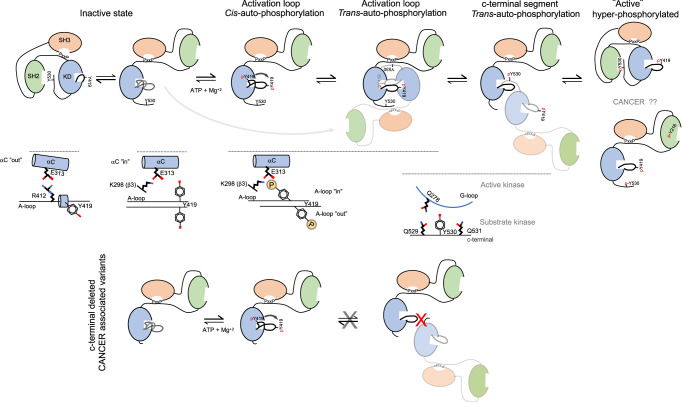
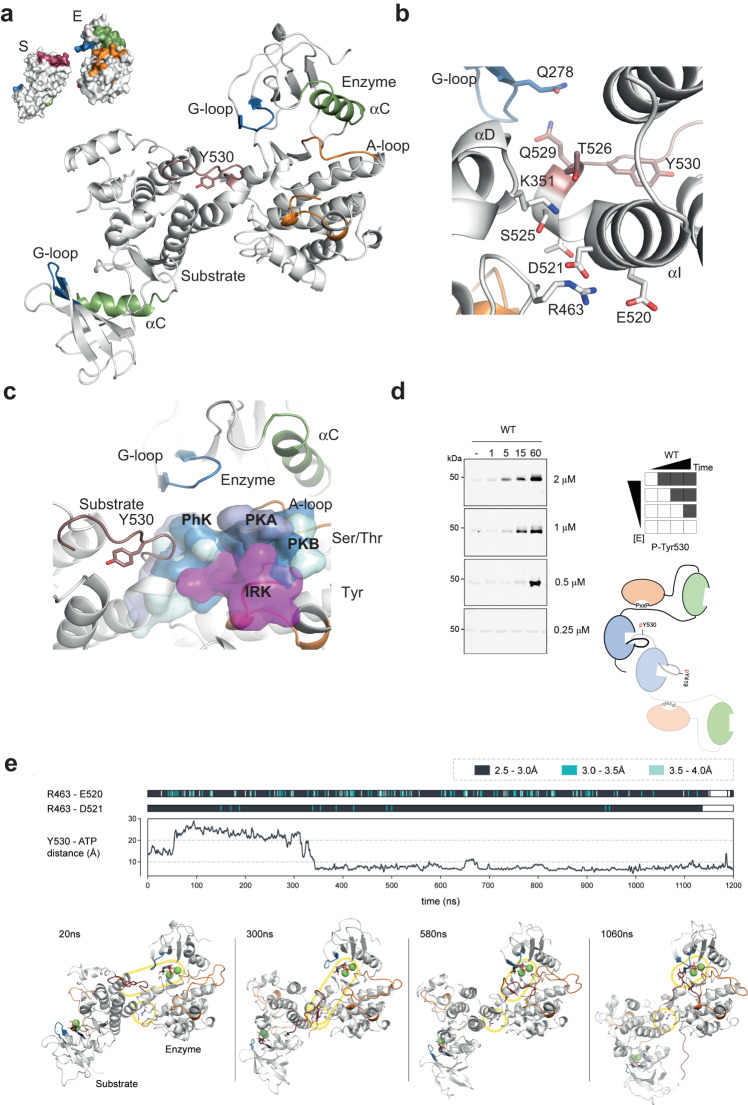
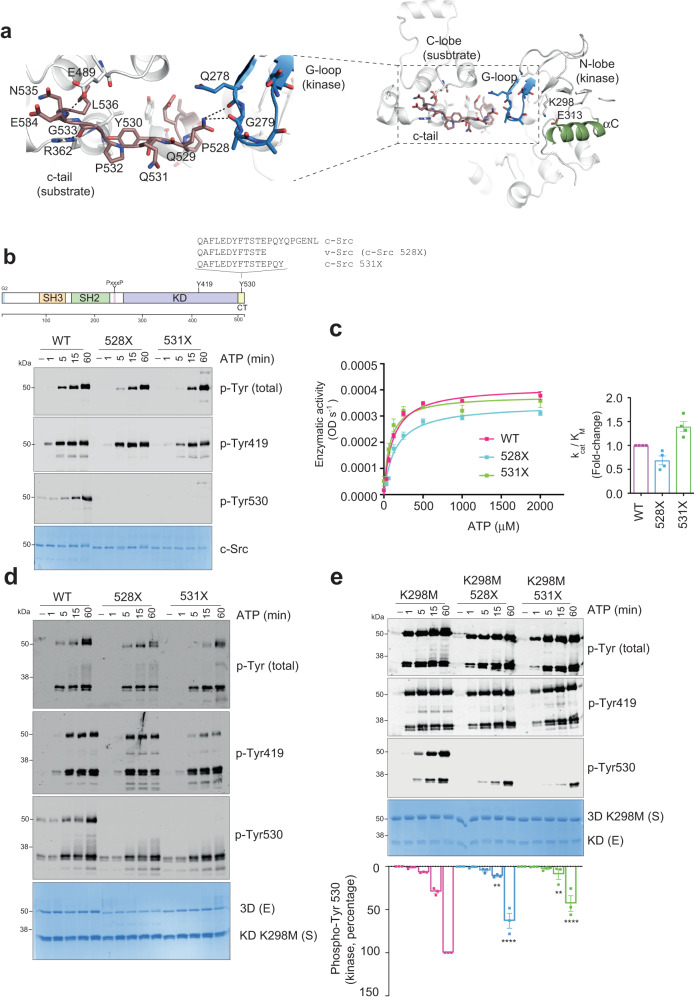
Autophosphorylation controls the transition between discrete functional and conformational states in protein kinases, yet the structural and molecular determinants underlying this fundamental process remain unclear. Here we show that c-terminal Tyr 530 is a de facto c-Src autophosphorylation site with slow time-resolution kinetics and a strong intermolecular component. On the contrary, activation-loop Tyr 419 undergoes faster kinetics and a cis-to-trans phosphorylation switch that controls c-terminal Tyr 530 autophosphorylation, enzyme specificity, and strikingly, c-Src non-catalytic function as a substrate. In line with this, we visualize by X-ray crystallography a snapshot of Tyr 530 intermolecular autophosphorylation. In an asymmetric arrangement of both catalytic domains, a c-terminal palindromic phospho-motif flanking Tyr 530 on the substrate molecule engages the G-loop of the active kinase adopting a position ready for entry into the catalytic cleft. Perturbation of the phospho-motif accounts for c-Src dysfunction as indicated by viral and colorectal cancer (CRC)-associated c-terminal deleted variants. We show that c-terminal residues 531 to 536 are required for c-Src Tyr 530 autophosphorylation, and such a detrimental effect is caused by the substrate molecule inhibiting allosterically the active kinase. Our work reveals a crosstalk between the activation and c-terminal segments that control the allosteric interplay between substrate- and enzyme-acting kinases during autophosphorylation.
自身磷酸化控制蛋白激酶在离散功能和构象状态之间的转变,但这一基本过程背后的结构和分子决定因素仍不清楚。在这里,我们表明 C 端 Tyr530 实际上是 c-Src 自身磷酸化位点,具有缓慢的时间分辨率动力学和强烈的分子间成分。相比之下,激活环 Tyr419 经历更快的动力学和顺式到反式磷酸化转换,控制 C 端 Tyr530 自身磷酸化、酶特异性,以及引人注目的 c-Src 非催化功能作为底物。与此一致,我们通过 X 射线晶体学可视化了 Tyr530 分子间自身磷酸化的瞬时快照。在两个催化结构域的不对称排列中,底物分子上 Tyr530 侧翼的 C 端回文磷酸化基序与活性激酶的 G 环结合,采用准备进入催化裂缝的位置。磷酸化基序的扰动解释了病毒和结直肠癌(CRC)相关 C 端缺失变体引起的 c-Src 功能障碍。我们表明,C 端残基 531 到 536 是 c-Src Tyr530 自身磷酸化所必需的,这种有害影响是由底物分子通过变构抑制活性激酶引起的。我们的工作揭示了激活和 C 端片段之间的串扰,控制了在自身磷酸化过程中底物和酶作用激酶之间的变构相互作用。