State Key Laboratory of Cellular Stress Biology, School of Life Sciences, Xiamen University, Xiamen, China.
Elife. 2024 Jan 17;13:e89615. doi: 10.7554/eLife.89615.
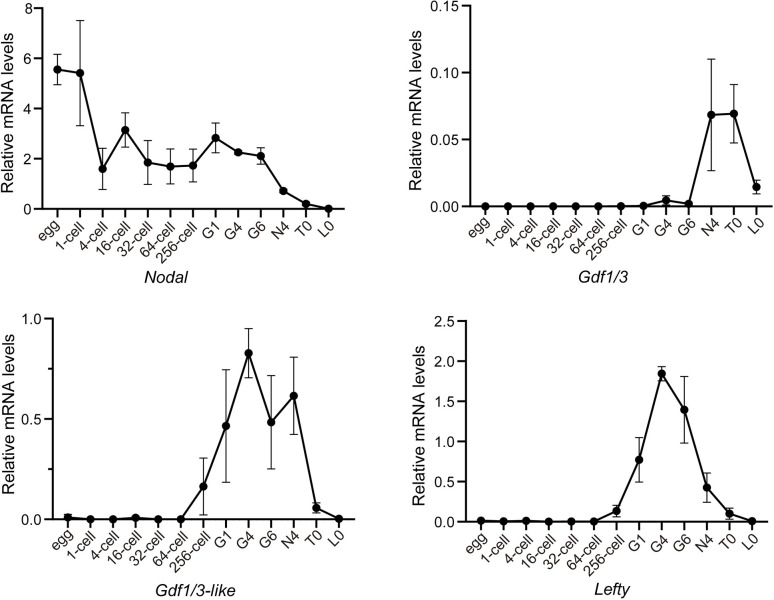
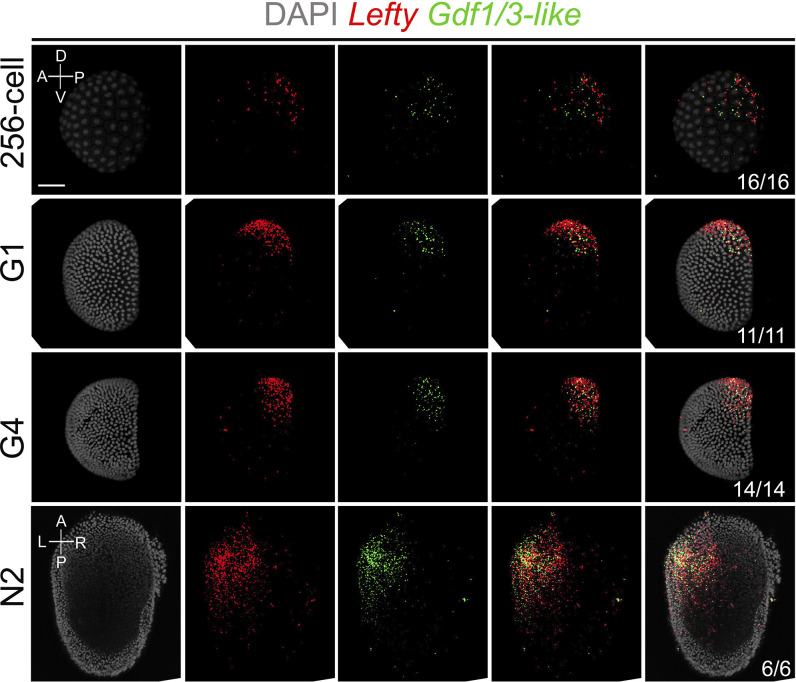
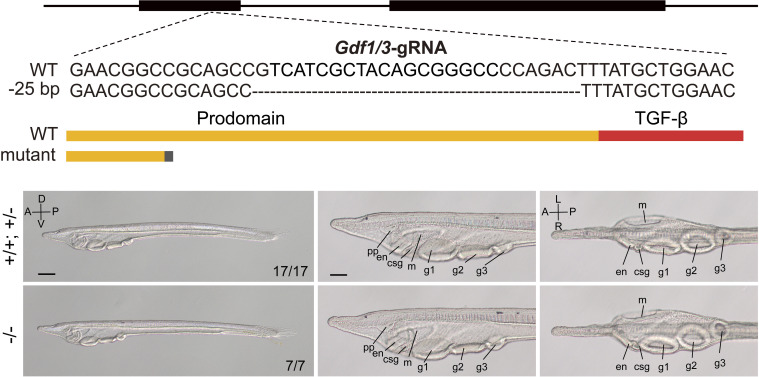
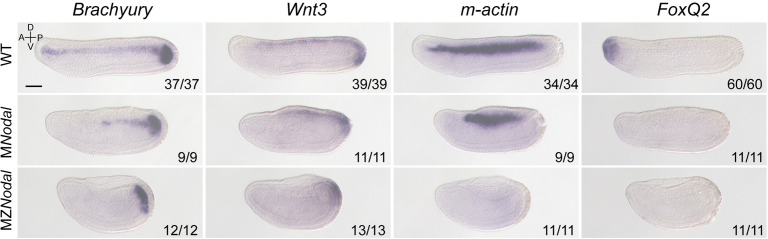
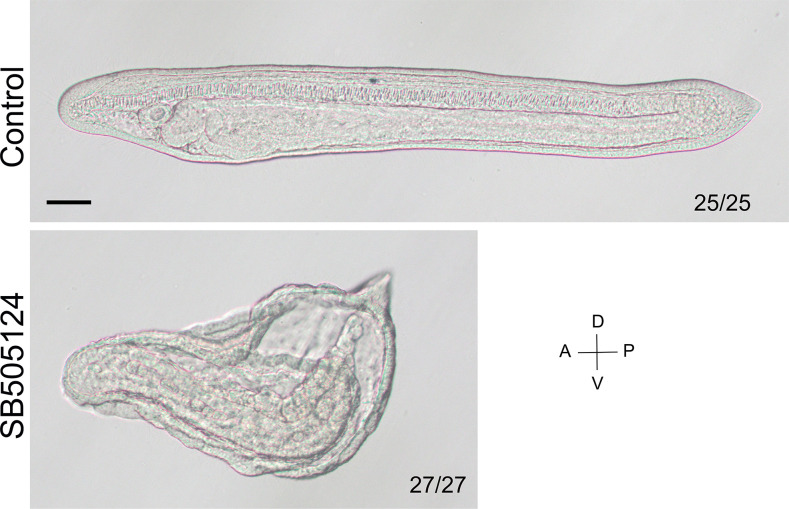
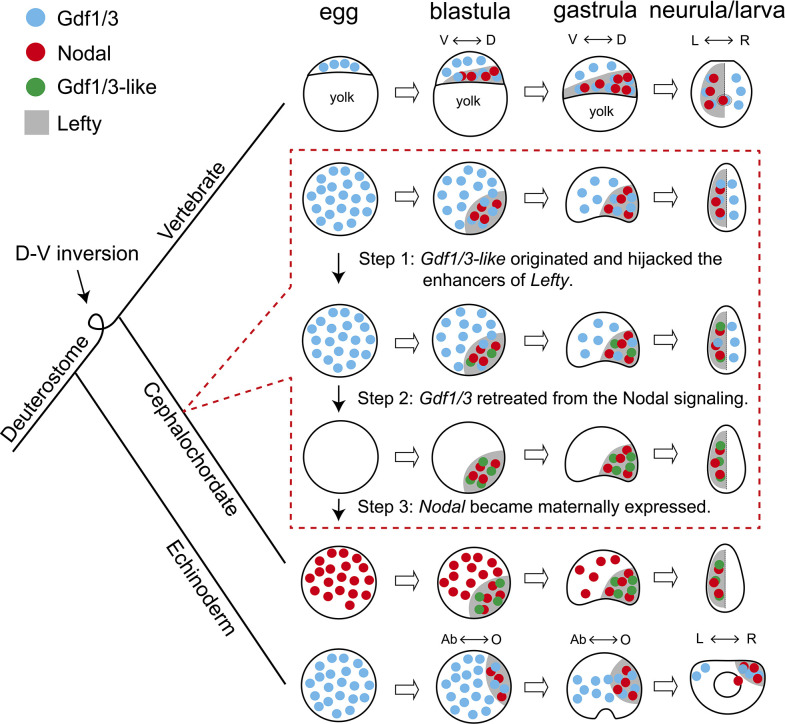
A central goal of evolutionary developmental biology is to decipher the evolutionary pattern of gene regulatory networks (GRNs) that control embryonic development, and the mechanism underlying GRNs evolution. The Nodal signaling that governs the body axes of deuterostomes exhibits a conserved GRN orchestrated principally by Nodal, Gdf1/3, and Lefty. Here we show that this GRN has been rewired in cephalochordate amphioxus. We found that while the amphioxus ortholog exhibited nearly no embryonic expression, its duplicate , linked to , was zygotically expressed in a similar pattern as . Consistent with this, while mutants showed defects in axial development, mutants did not. Further transgenic analyses showed that the intergenic region between and could drive reporter gene expression as that of the two genes. These results indicated that has taken over the axial development role of in amphioxus, possibly through hijacking enhancers. We finally demonstrated that, to compensate for the loss of maternal expression, Nodal has become an indispensable maternal factor in amphioxus and its maternal mutants caused axial defects as mutants. We therefore demonstrated a case that the evolution of GRNs could be triggered by enhancer hijacking events. This pivotal event has allowed the emergence of a new GRN in extant amphioxus, presumably through a stepwise process. In addition, the co-expression of and achieved by a shared regulatory region may have provided robustness during body axis formation, which provides a selection-based hypothesis for the phenomena called developmental system drift.
进化发育生物学的一个核心目标是破译控制胚胎发育的基因调控网络 (GRN) 的进化模式及其进化的机制。调控后口动物体轴的 Nodal 信号表现出主要由 Nodal、Gdf1/3 和 Lefty 协调的保守 GRN。在这里,我们表明这种 GRN 在文昌鱼中有重布线。我们发现,虽然文昌鱼的同源物几乎没有胚胎表达,但它的副本,与,在类似的模式中合子表达,与。与此一致的是,虽然突变体显示出轴向发育缺陷,但突变体没有。进一步的转基因分析表明,和之间的基因间区可以驱动报告基因的表达,就像两个基因一样。这些结果表明,在文昌鱼中,已经接管了的轴向发育作用,可能是通过劫持的增强子。我们最后证明,为了弥补母本表达的丧失,Nodal 在文昌鱼中已成为不可或缺的母本因子,其母本突变体引起轴向缺陷,如突变体。因此,我们证明了 GRN 的进化可以通过增强子劫持事件引发。这一关键事件允许新的 GRN 在现存的文昌鱼中出现,可能是通过一个逐步的过程。此外,通过共享调控区实现的和的共表达可能在体轴形成过程中提供了稳健性,这为称为发育系统漂移的现象提供了一个基于选择的假设。