Molecular Pathology Section, Laboratory of Immunogenetics, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Bethesda, MD, 20892, USA.
Center for Cancer and Immunology Research, Children's National Research Institute, Washington, DC, 20010, USA.
Genome Biol. 2024 Jan 31;25(1):40. doi: 10.1186/s13059-024-03175-0.
Pervasive usage of alternative promoters leads to the deregulation of gene expression in carcinogenesis and may drive the emergence of new genes in spermatogenesis. However, little is known regarding the mechanisms underpinning the activation of alternative promoters.
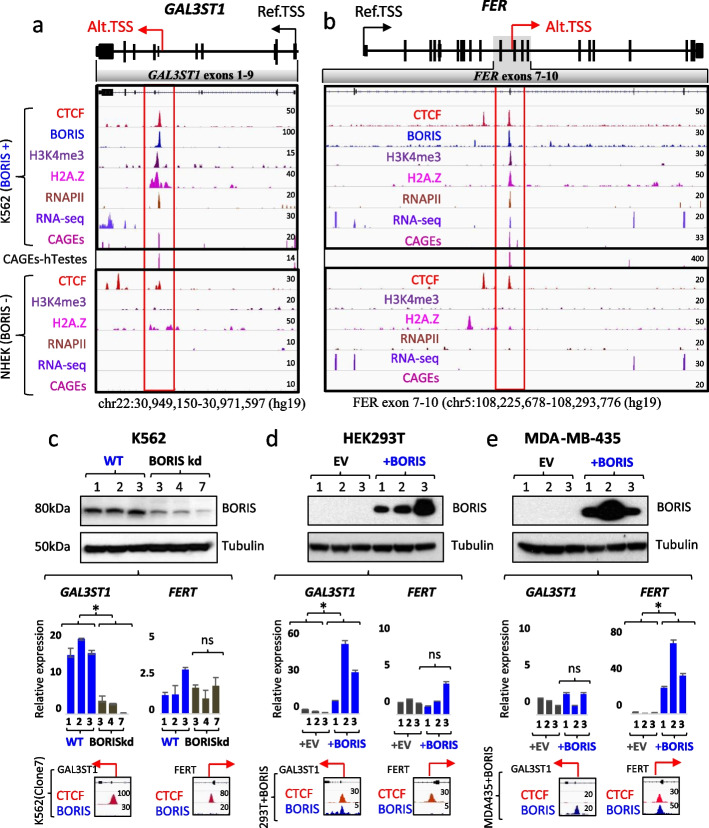
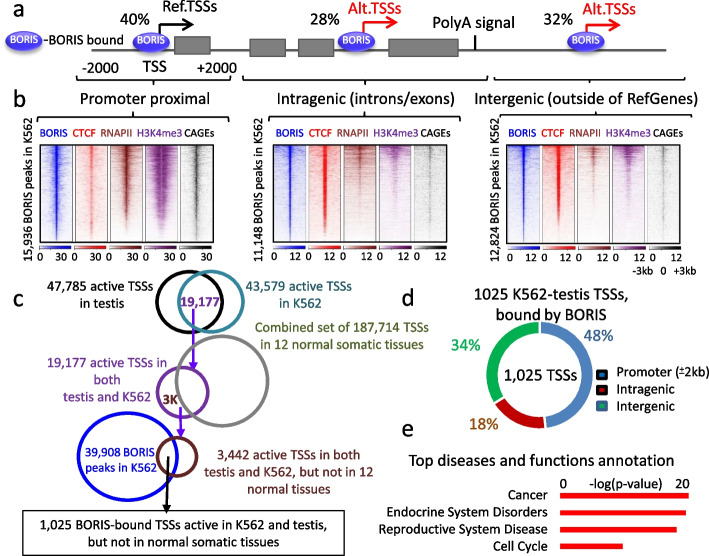
Here we describe how alternative cancer-testis-specific transcription is activated. We show that intergenic and intronic CTCF binding sites, which are transcriptionally inert in normal somatic cells, could be epigenetically reprogrammed into active de novo promoters in germ and cancer cells. BORIS/CTCFL, the testis-specific paralog of the ubiquitously expressed CTCF, triggers the epigenetic reprogramming of CTCF sites into units of active transcription. BORIS binding initiates the recruitment of the chromatin remodeling factor, SRCAP, followed by the replacement of H2A histone with H2A.Z, resulting in a more relaxed chromatin state in the nucleosomes flanking the CTCF binding sites. The relaxation of chromatin around CTCF binding sites facilitates the recruitment of multiple additional transcription factors, thereby activating transcription from a given binding site. We demonstrate that the epigenetically reprogrammed CTCF binding sites can drive the expression of cancer-testis genes, long noncoding RNAs, retro-pseudogenes, and dormant transposable elements.
Thus, BORIS functions as a transcription factor that epigenetically reprograms clustered CTCF binding sites into transcriptional start sites, promoting transcription from alternative promoters in both germ cells and cancer cells.
普遍存在的替代启动子的使用导致了致癌过程中基因表达的失调,并可能在精子发生中驱动新基因的出现。然而,对于支持替代启动子激活的机制知之甚少。
在这里,我们描述了如何激活替代性癌症-睾丸特异性转录。我们表明,在正常体细胞中转录沉默的基因间和内含子 CTCF 结合位点,在生殖细胞和癌细胞中可以通过表观遗传重新编程为新的活性启动子。BORIS/CTCFL 是普遍表达的 CTCF 的睾丸特异性同源物,触发了 CTCF 位点向活性转录单位的表观遗传重编程。BORIS 结合引发了染色质重塑因子 SRCAP 的招募,随后用 H2A.Z 取代 H2A 组蛋白,导致围绕 CTCF 结合位点的核小体中的染色质状态更为松弛。染色质在 CTCF 结合位点周围的松弛有助于多个额外转录因子的招募,从而激活给定结合位点的转录。我们证明,表观遗传重编程的 CTCF 结合位点可以驱动癌症-睾丸基因、长非编码 RNA、反转录假基因和休眠转座元件的表达。
因此,BORIS 作为一种转录因子,通过表观遗传将簇集的 CTCF 结合位点重新编程为转录起始位点,促进生殖细胞和癌细胞中替代启动子的转录。