Department of Pathology, New York University Langone Health, New York, NY, 10016, USA.
Gladstone Institutes, San Francisco, CA, 94158, USA.
Genome Biol. 2020 May 11;21(1):108. doi: 10.1186/s13059-020-02024-0.
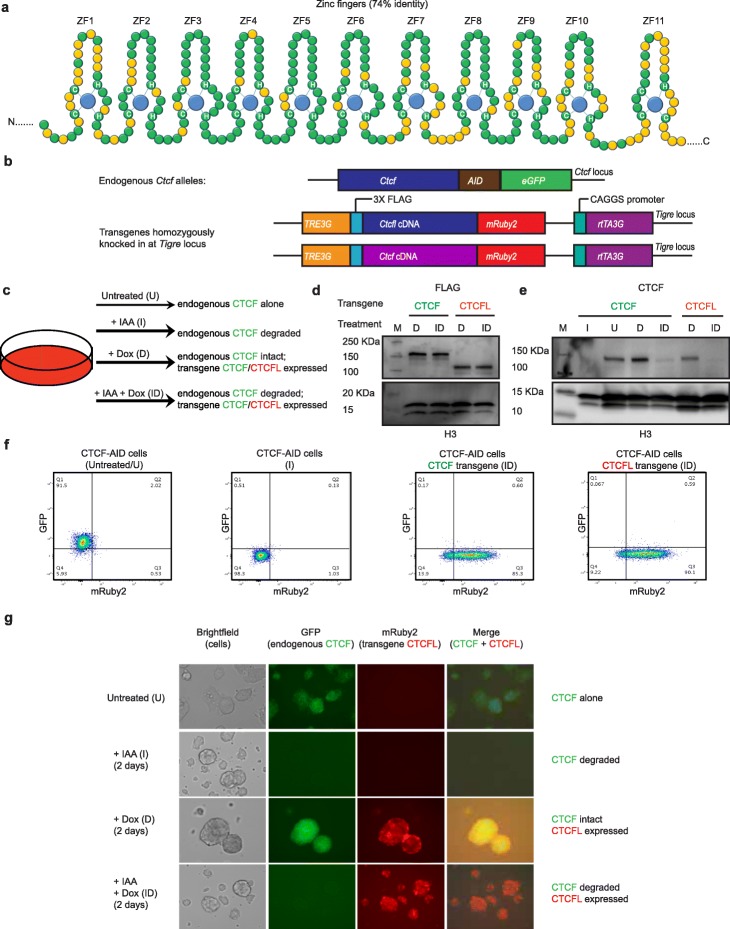
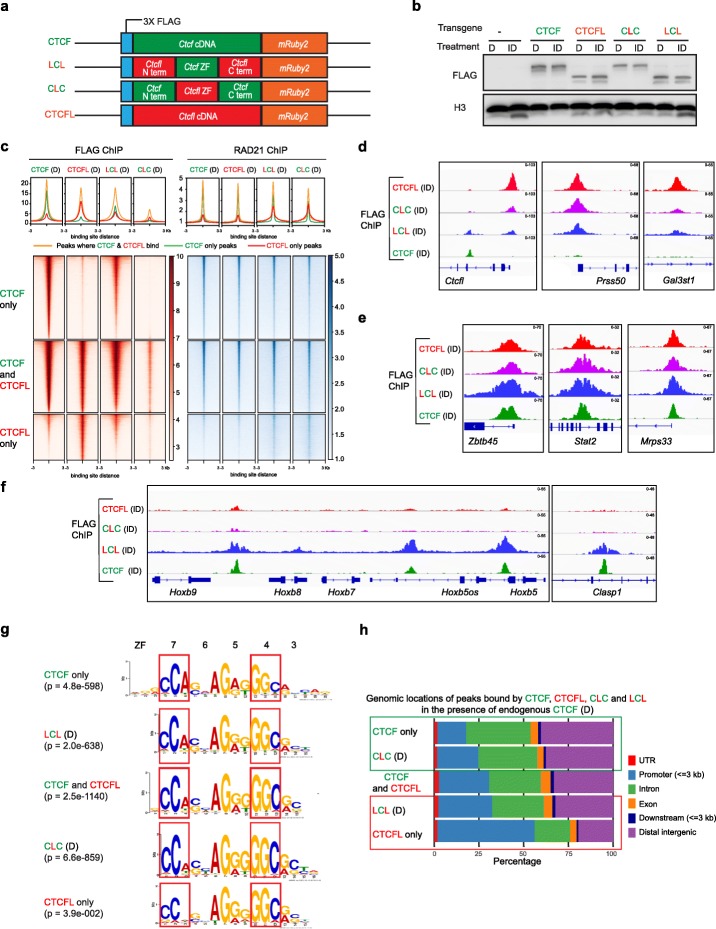
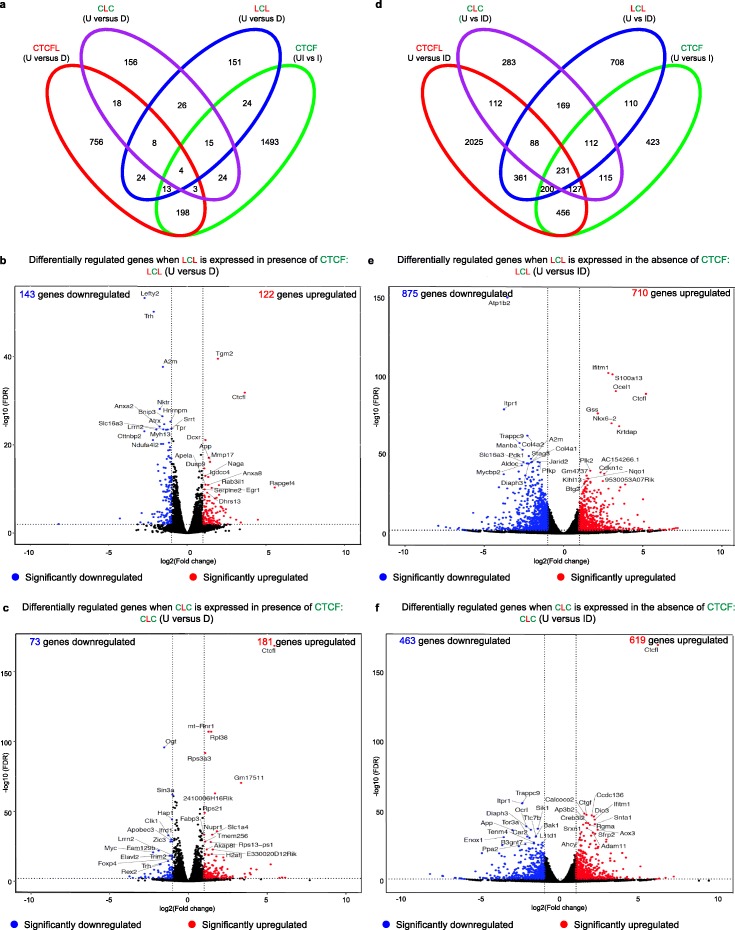
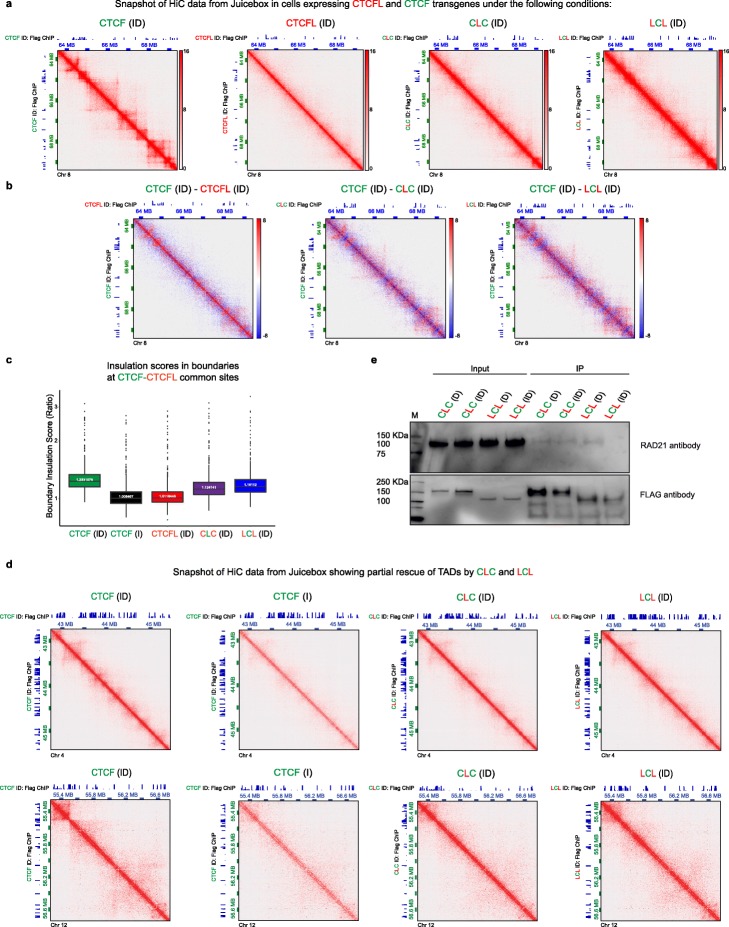
Ubiquitously expressed CTCF is involved in numerous cellular functions, such as organizing chromatin into TAD structures. In contrast, its paralog, CTCFL, is normally only present in the testis. However, it is also aberrantly expressed in many cancers. While it is known that shared and unique zinc finger sequences in CTCF and CTCFL enable CTCFL to bind competitively to a subset of CTCF binding sites as well as its own unique locations, the impact of CTCFL on chromosome organization and gene expression has not been comprehensively analyzed in the context of CTCF function. Using an inducible complementation system, we analyze the impact of expressing CTCFL and CTCF-CTCFL chimeric proteins in the presence or absence of endogenous CTCF to clarify the relative and combined contribution of CTCF and CTCFL to chromosome organization and transcription.
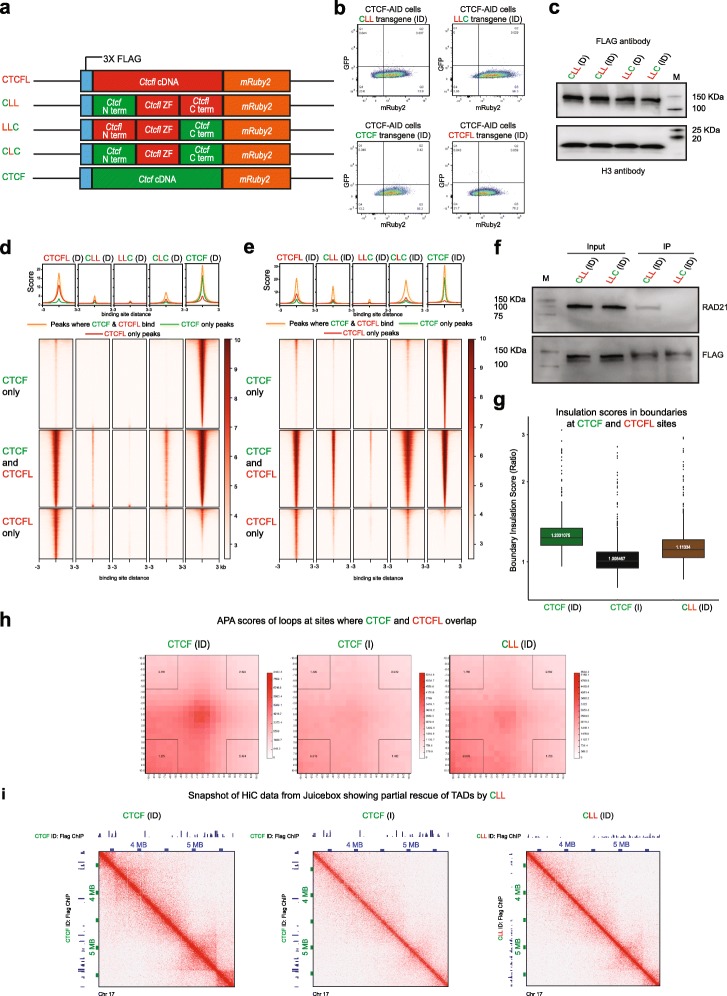
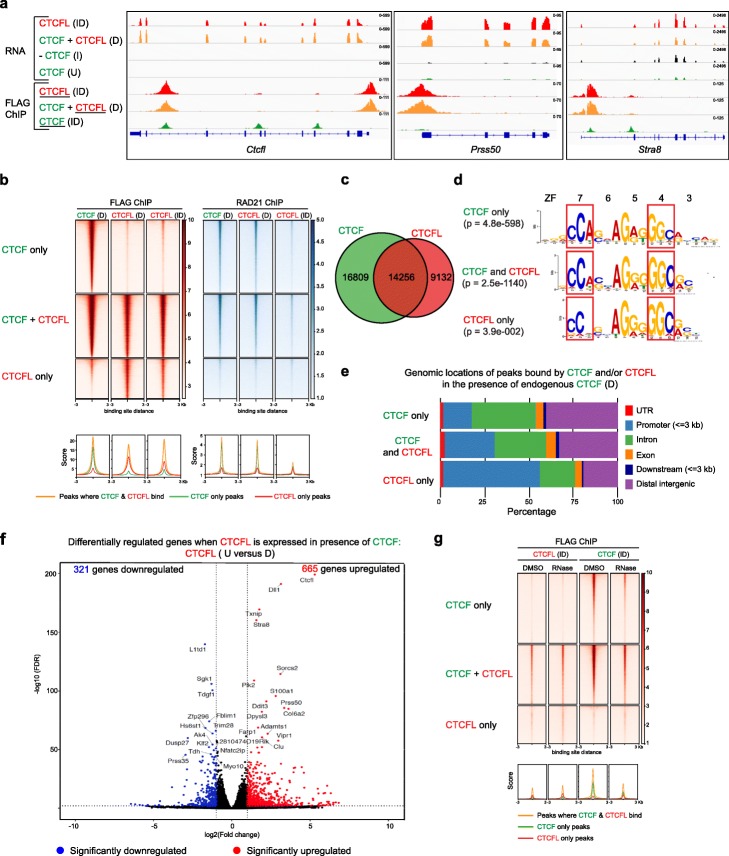
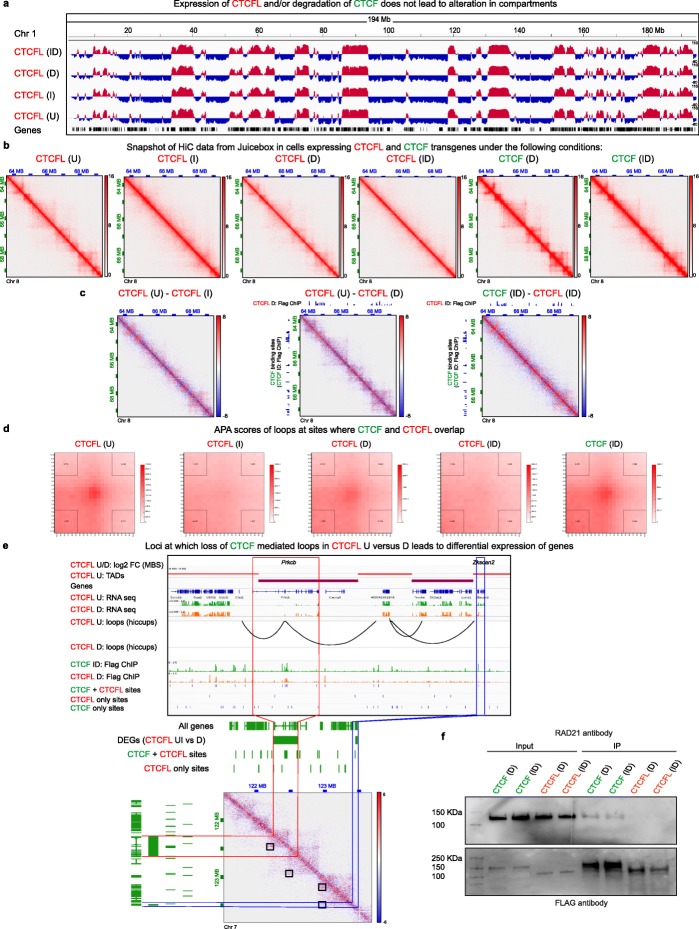
We demonstrate that the N terminus of CTCF interacts with cohesin which explains the requirement for convergent CTCF binding sites in loop formation. By analyzing CTCF and CTCFL binding in tandem, we identify phenotypically distinct sites with respect to motifs, targeting to promoter/intronic intergenic regions and chromatin folding. Finally, we reveal that the N, C, and zinc finger terminal domains play unique roles in targeting each paralog to distinct binding sites to regulate transcription, chromatin looping, and insulation.
This study clarifies the unique and combined contribution of CTCF and CTCFL to chromosome organization and transcription, with direct implications for understanding how their co-expression deregulates transcription in cancer.
普遍表达的 CTCF 参与许多细胞功能,如将染色质组织成 TAD 结构。相比之下,其同源物 CTCFL 通常仅存在于睾丸中。然而,它也在许多癌症中异常表达。虽然已知 CTCF 和 CTCFL 中共享和独特的锌指序列使 CTCFL 能够与 CTCF 结合位点的子集以及其自身独特的位置竞争结合,但 CTCFL 对染色体组织和基因表达的影响尚未在 CTCF 功能的背景下进行全面分析。使用可诱导的互补系统,我们分析了在存在或不存在内源性 CTCF 的情况下表达 CTCFL 和 CTCF-CTCFL 嵌合蛋白的影响,以阐明 CTCF 和 CTCFL 对染色体组织和转录的相对和综合贡献。
我们证明 CTCF 的 N 端与 cohesin 相互作用,这解释了环形成中需要会聚的 CTCF 结合位点。通过分析串联的 CTCF 和 CTCFL 结合,我们确定了在基序、靶向启动子/内含子基因间区和染色质折叠方面具有表型不同的位点。最后,我们揭示 N、C 和锌指末端结构域在将每个同源物靶向到不同的结合位点以调节转录、染色质环化和隔离方面发挥独特的作用。
这项研究阐明了 CTCF 和 CTCFL 对染色体组织和转录的独特和综合贡献,对理解它们的共表达如何在癌症中失调转录具有直接意义。