Han Hui, Luo Rong-Hua, Long Xin-Yan, Wang Li-Qiong, Zhu Qian, Tang Xin-Yue, Zhu Rui, Ma Yi-Cheng, Zheng Yong-Tang, Zou Cheng-Gang
State Key Laboratory for Conservation and Utilization of Bio-Resources in Yunnan, School of Life Sciences, Yunnan University, Kunming, China.
Key Laboratory of Bioactive Peptides of Yunnan Province/Key Laboratory of Animal Models and Human Disease Mechanisms of the Chinese Academy of Sciences, KIZ-CUHK Joint Laboratory of Bio-Resources and Molecular Research in Common Diseases, Center for Biosafety Mega-Science, Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, China.
Elife. 2024 Feb 20;13:e85985. doi: 10.7554/eLife.85985.
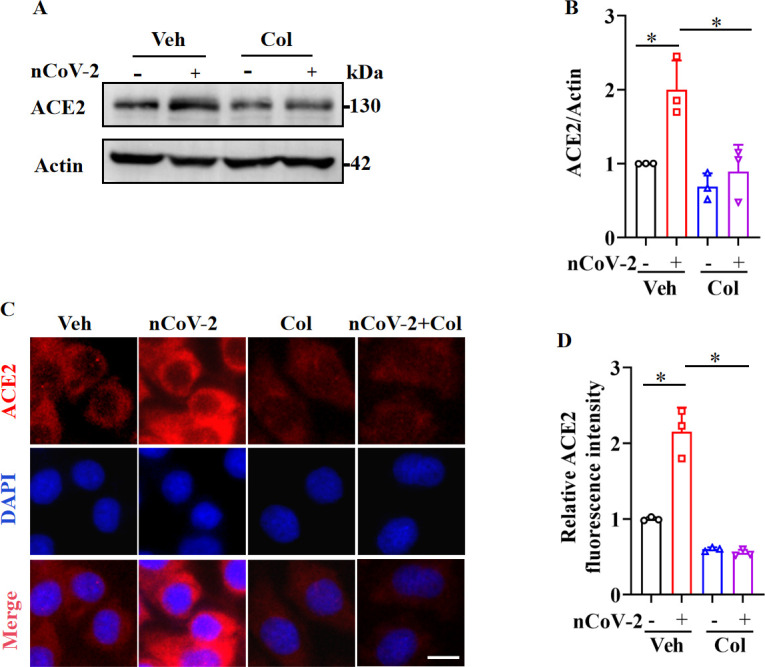
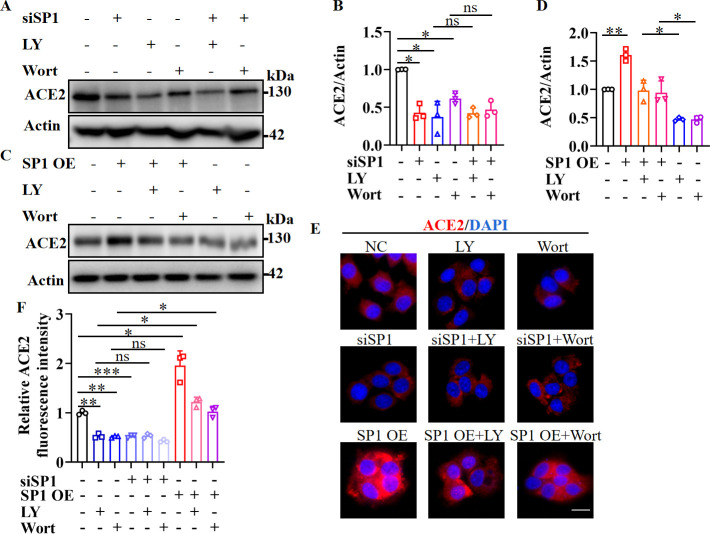
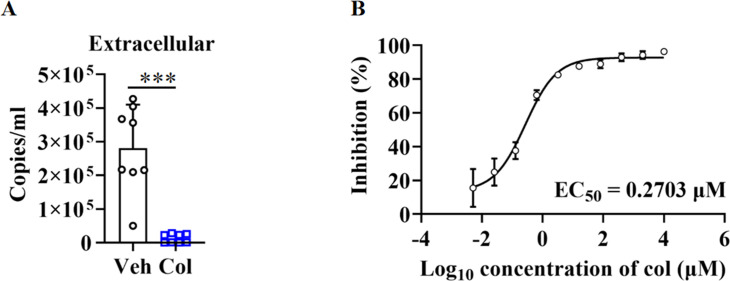
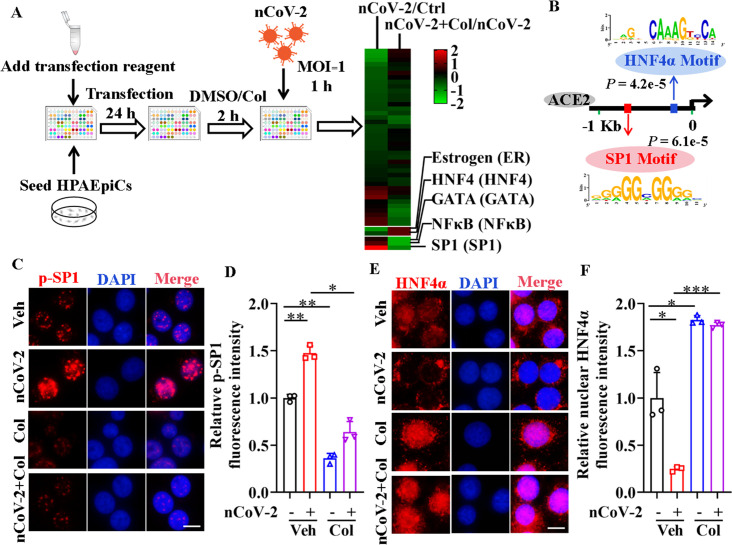
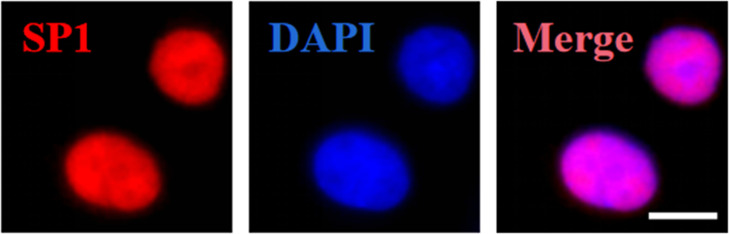
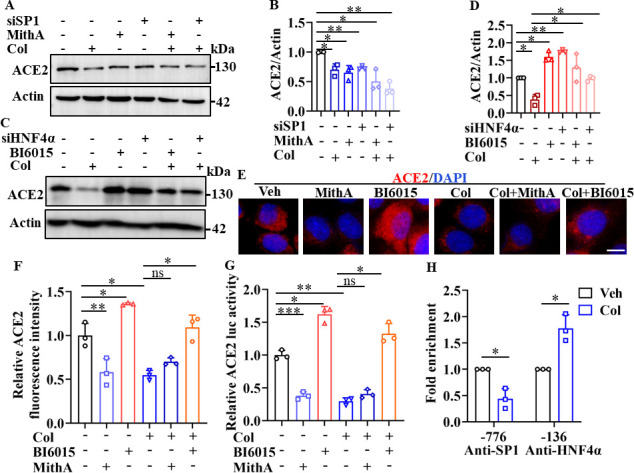
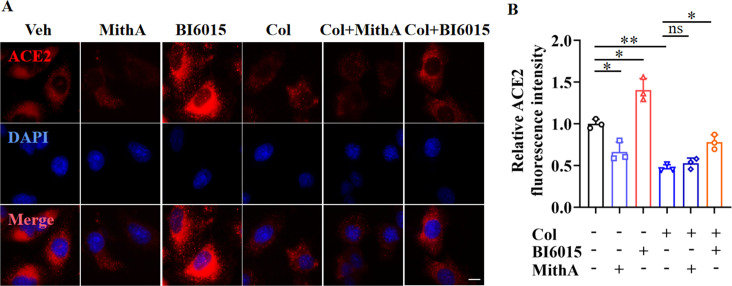
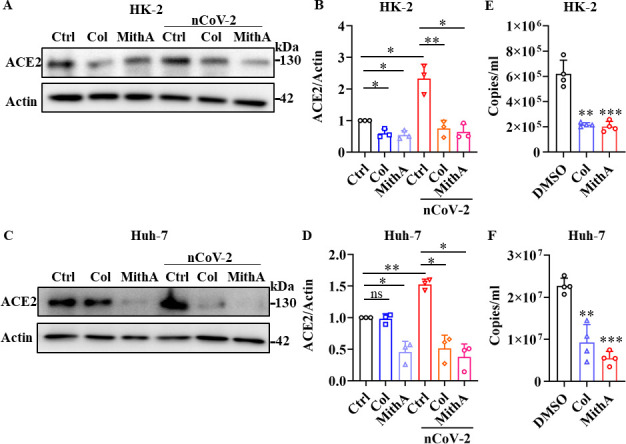
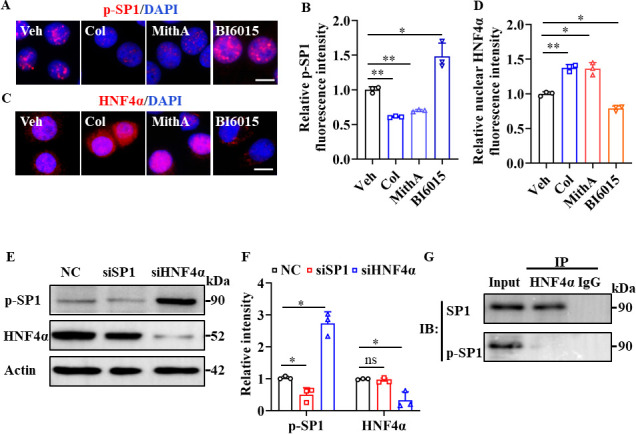
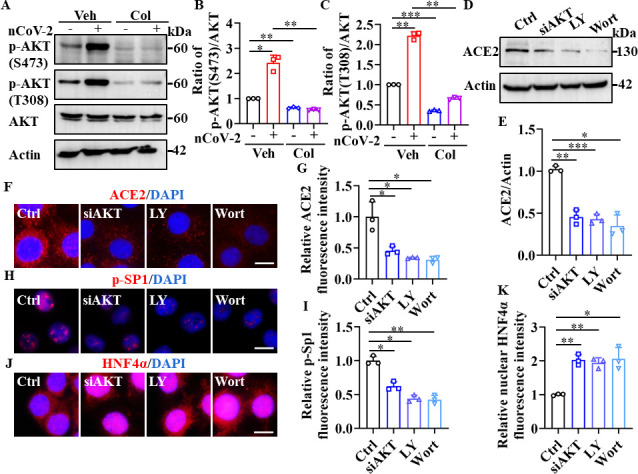
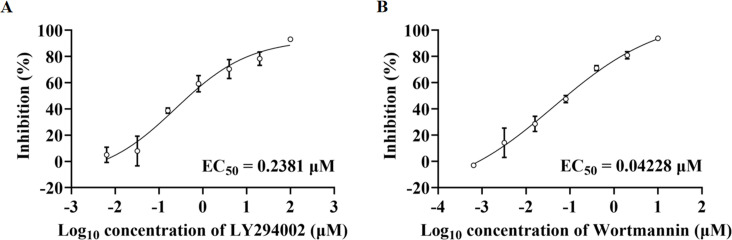
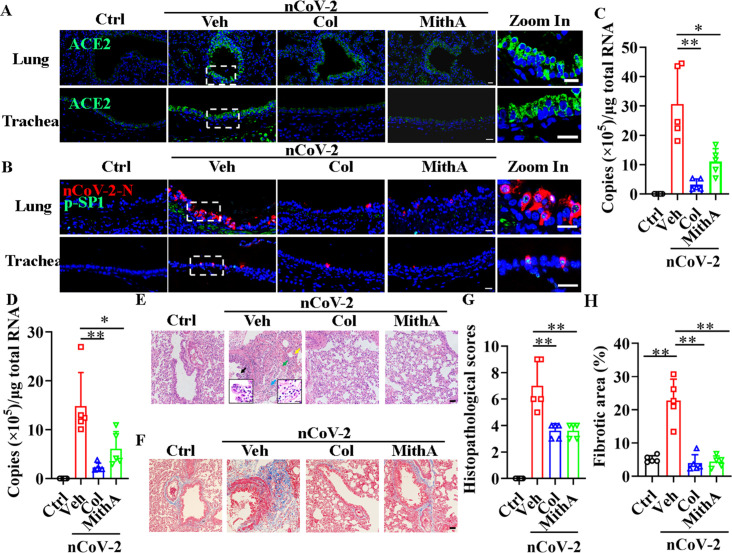
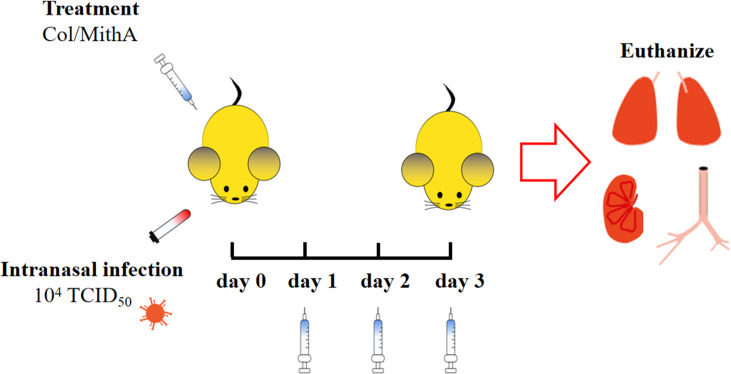
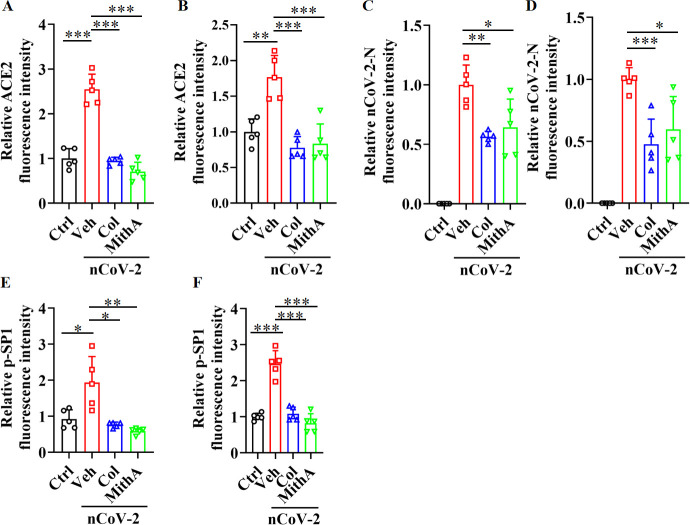
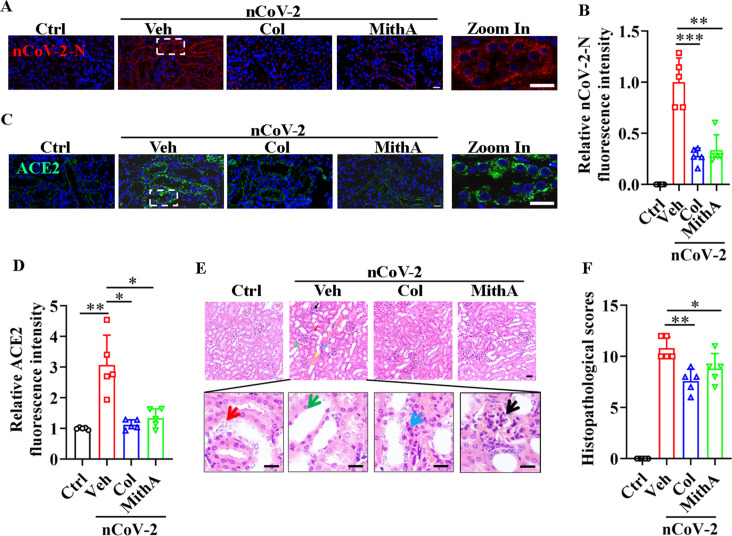
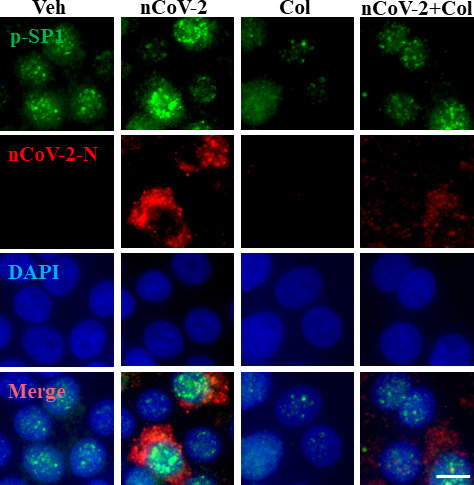
Angiotensin-converting enzyme 2 (ACE2) is a major cell entry receptor for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). The induction of ACE2 expression may serve as a strategy by SARS-CoV-2 to facilitate its propagation. However, the regulatory mechanisms of ACE2 expression after viral infection remain largely unknown. Using 45 different luciferase reporters, the transcription factors SP1 and HNF4α were found to positively and negatively regulate ACE2 expression, respectively, at the transcriptional level in human lung epithelial cells (HPAEpiCs). SARS-CoV-2 infection increased the transcriptional activity of SP1 while inhibiting that of HNF4α. The PI3K/AKT signaling pathway, activated by SARS-CoV-2 infection, served as a crucial regulatory node, inducing ACE2 expression by enhancing SP1 phosphorylation-a marker of its activity-and reducing the nuclear localization of HNF4α. However, colchicine treatment inhibited the PI3K/AKT signaling pathway, thereby suppressing ACE2 expression. In Syrian hamsters () infected with SARS-CoV-2, inhibition of SP1 by either mithramycin A or colchicine resulted in reduced viral replication and tissue injury. In summary, our study uncovers a novel function of SP1 in the regulation of ACE2 expression and identifies SP1 as a potential target to reduce SARS-CoV-2 infection.
血管紧张素转换酶2(ACE2)是严重急性呼吸综合征冠状病毒2(SARS-CoV-2)的主要细胞进入受体。诱导ACE2表达可能是SARS-CoV-2促进其传播的一种策略。然而,病毒感染后ACE2表达的调控机制仍 largely未知。使用45种不同的荧光素酶报告基因,发现转录因子SP1和HNF4α在人肺上皮细胞(HPAEpiCs)中分别在转录水平上正向和负向调节ACE2表达。SARS-CoV-2感染增加了SP1的转录活性,同时抑制了HNF4α的转录活性。由SARS-CoV-2感染激活的PI3K/AKT信号通路作为一个关键的调控节点,通过增强SP1磷酸化(其活性的标志物)并减少HNF4α的核定位来诱导ACE2表达。然而,秋水仙碱处理抑制了PI3K/AKT信号通路,从而抑制了ACE2表达。在感染SARS-CoV-2的叙利亚仓鼠中,放线菌素A或秋水仙碱对SP1的抑制导致病毒复制和组织损伤减少。总之,我们的研究揭示了SP1在调节ACE2表达中的新功能,并将SP1确定为减少SARS-CoV-2感染的潜在靶点。