Department of Biology, Philipps-University Marburg, Marburg, Germany.
Department of Molecular and Cell Biology and Howard Hughes Medical Institute, University of California, Berkeley, California, United States of America.
PLoS Biol. 2024 Feb 20;22(2):e3002508. doi: 10.1371/journal.pbio.3002508. eCollection 2024 Feb.
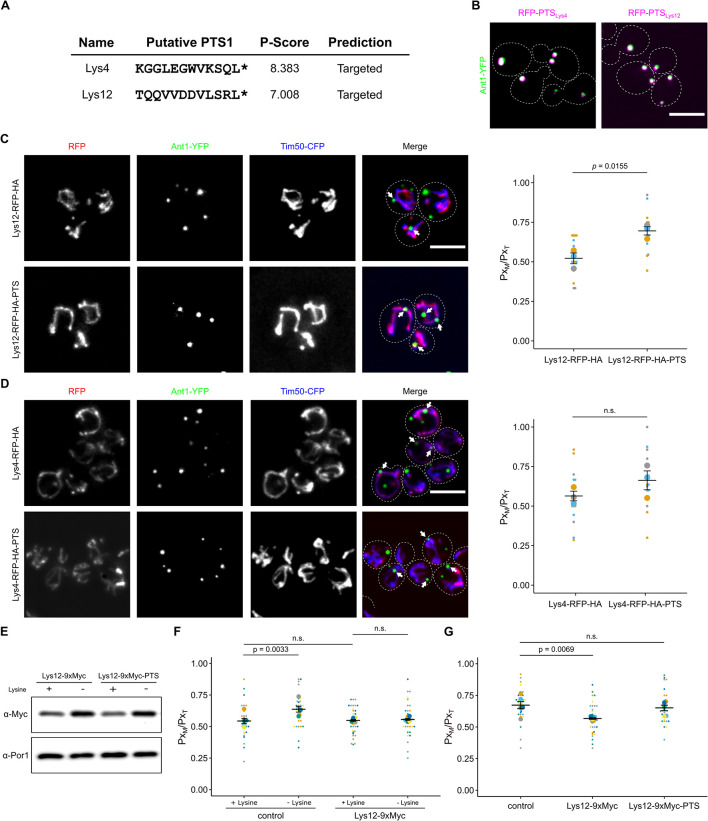
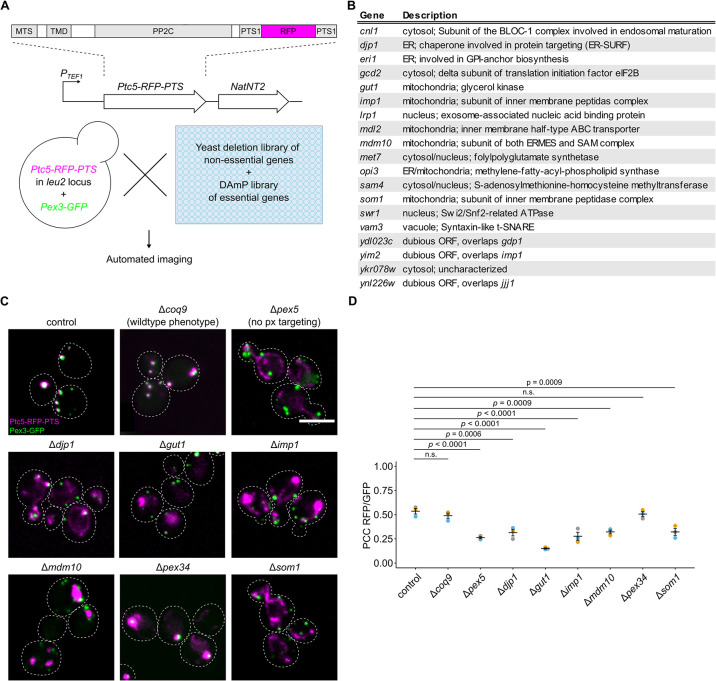
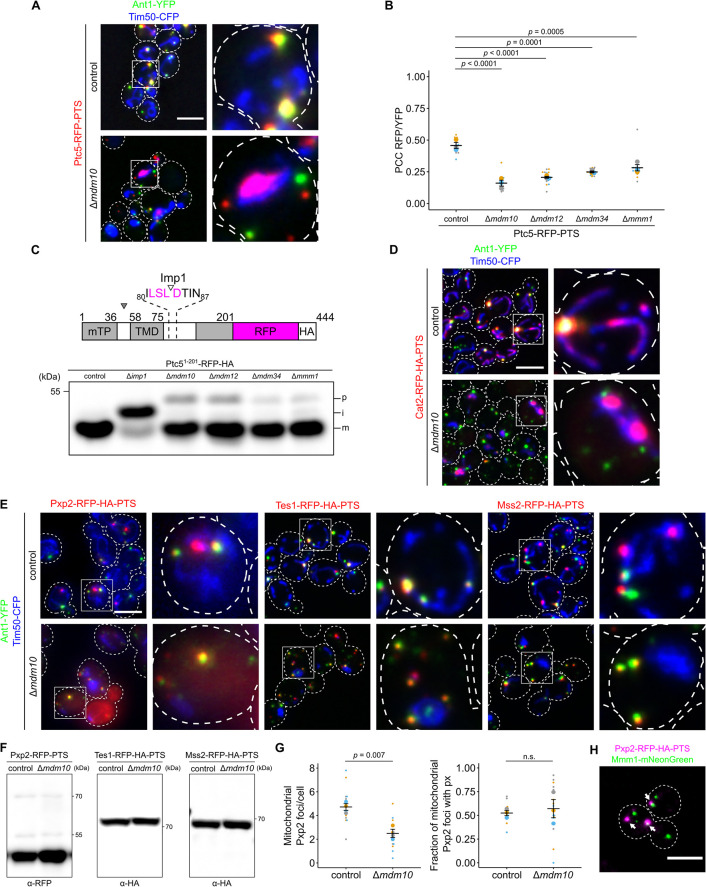
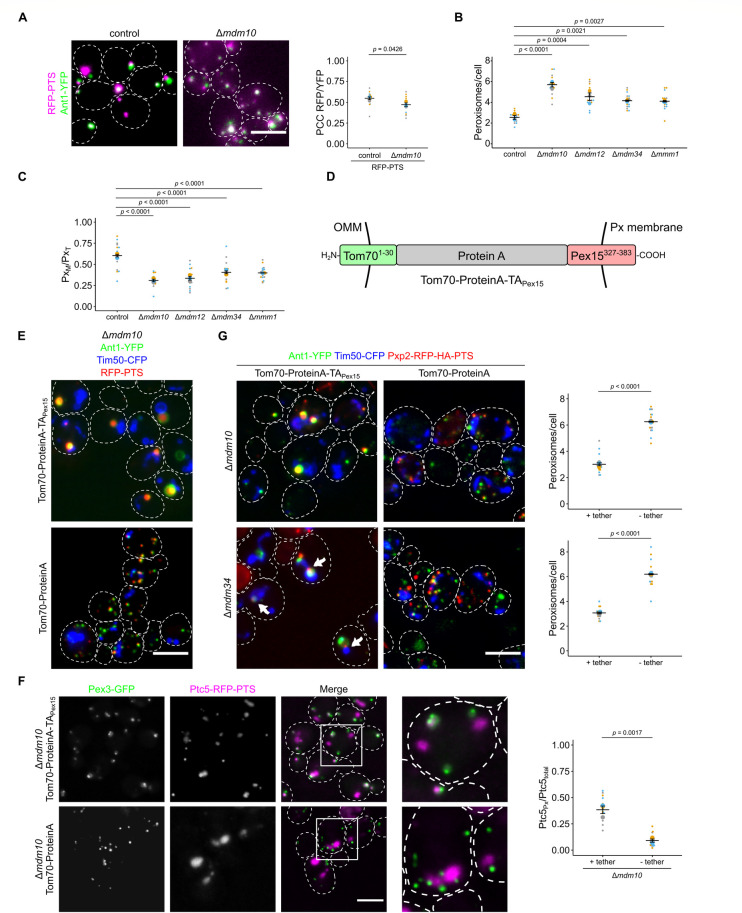
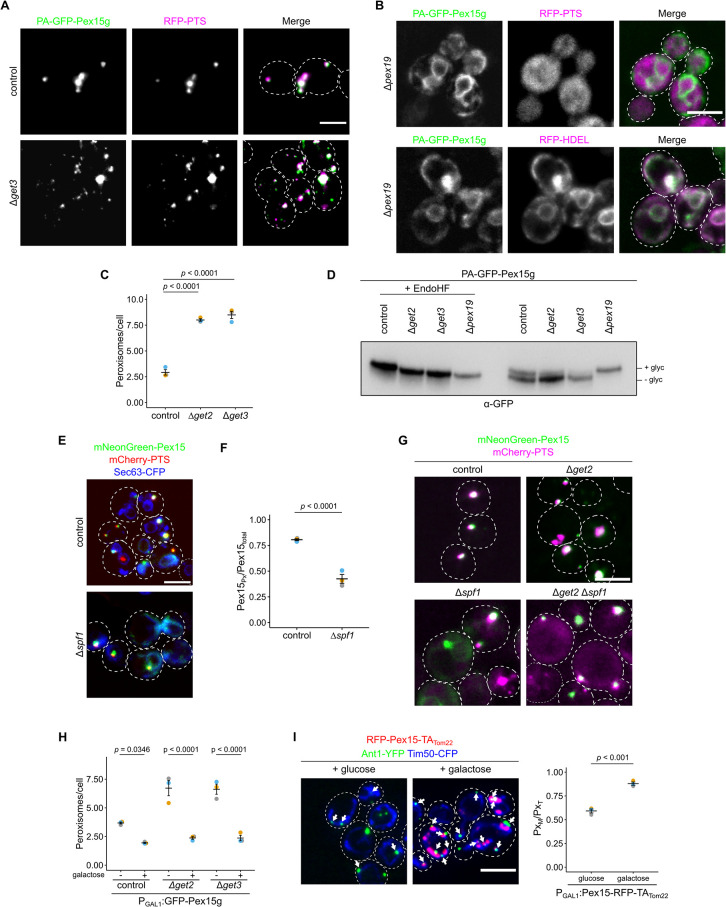
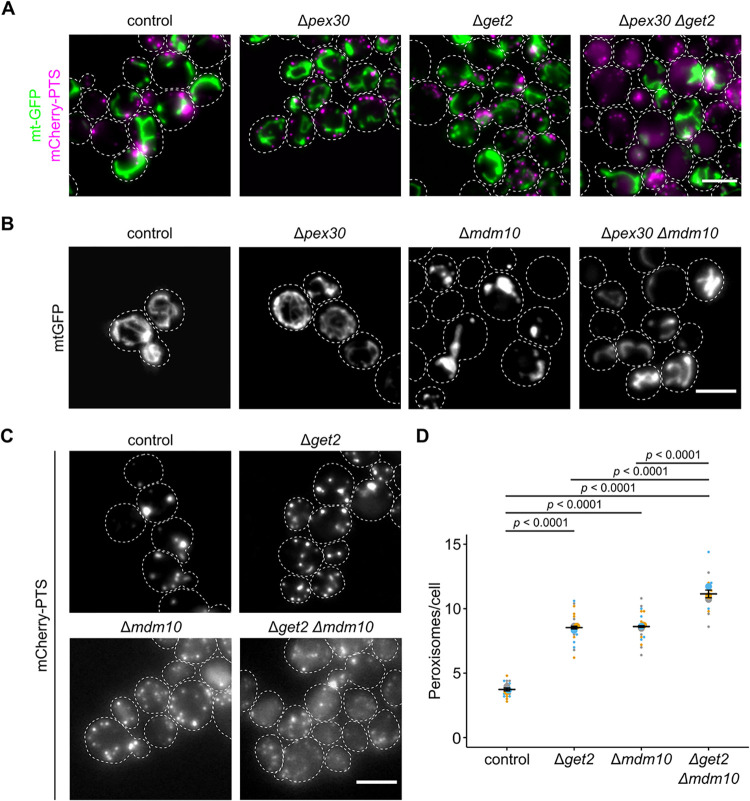
Peroxisomes are organelles with crucial functions in oxidative metabolism. To correctly target to peroxisomes, proteins require specialized targeting signals. A mystery in the field is the sorting of proteins that carry a targeting signal for peroxisomes and as well as for other organelles, such as mitochondria or the endoplasmic reticulum (ER). Exploring several of these proteins in fungal model systems, we observed that they can act as tethers bridging organelles together to create contact sites. We show that in Saccharomyces cerevisiae this mode of tethering involves the peroxisome import machinery, the ER-mitochondria encounter structure (ERMES) at mitochondria and the guided entry of tail-anchored proteins (GET) pathway at the ER. Our findings introduce a previously unexplored concept of how dual affinity proteins can regulate organelle attachment and communication.
过氧化物酶体是具有氧化代谢关键功能的细胞器。为了正确靶向过氧化物酶体,蛋白质需要专门的靶向信号。该领域的一个谜团是对携带过氧化物酶体和其他细胞器(如线粒体或内质网(ER))靶向信号的蛋白质进行分类。通过在真菌模型系统中探索几种这些蛋白质,我们观察到它们可以作为连接细胞器的系链,以形成接触位点。我们表明,在酿酒酵母中,这种系链模式涉及过氧化物酶体导入机制、线粒体上的内质网-线粒体接触结构 (ERMES) 和内质网上的引导进入的尾部锚定蛋白 (GET) 途径。我们的发现引入了一个以前未被探索的概念,即双重亲和力蛋白如何调节细胞器的附着和通讯。