Department of Neuroscience, University of Copenhagen, Copenhagen, Denmark.
Heidelberg University Biochemistry Center, Heidelberg, Germany.
Elife. 2024 Feb 27;12:RP88619. doi: 10.7554/eLife.88619.
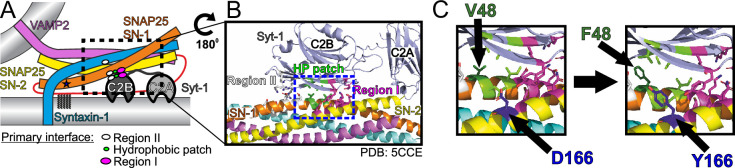
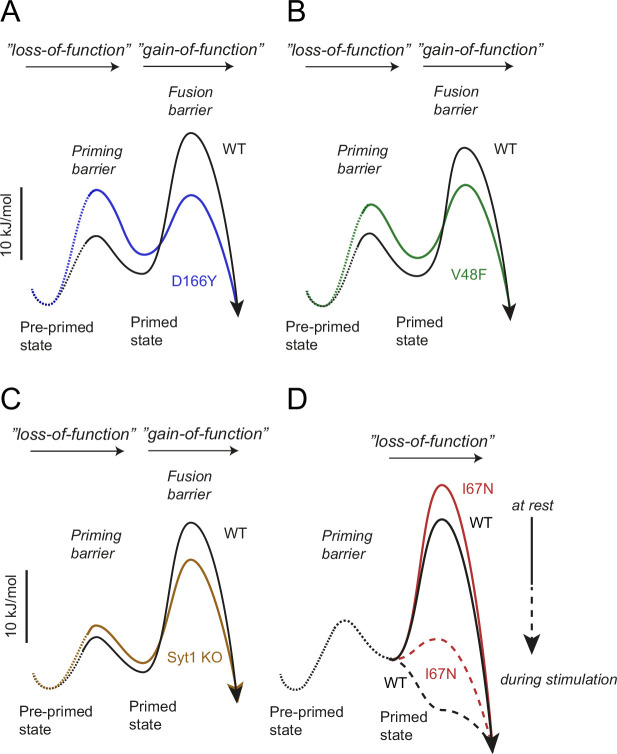
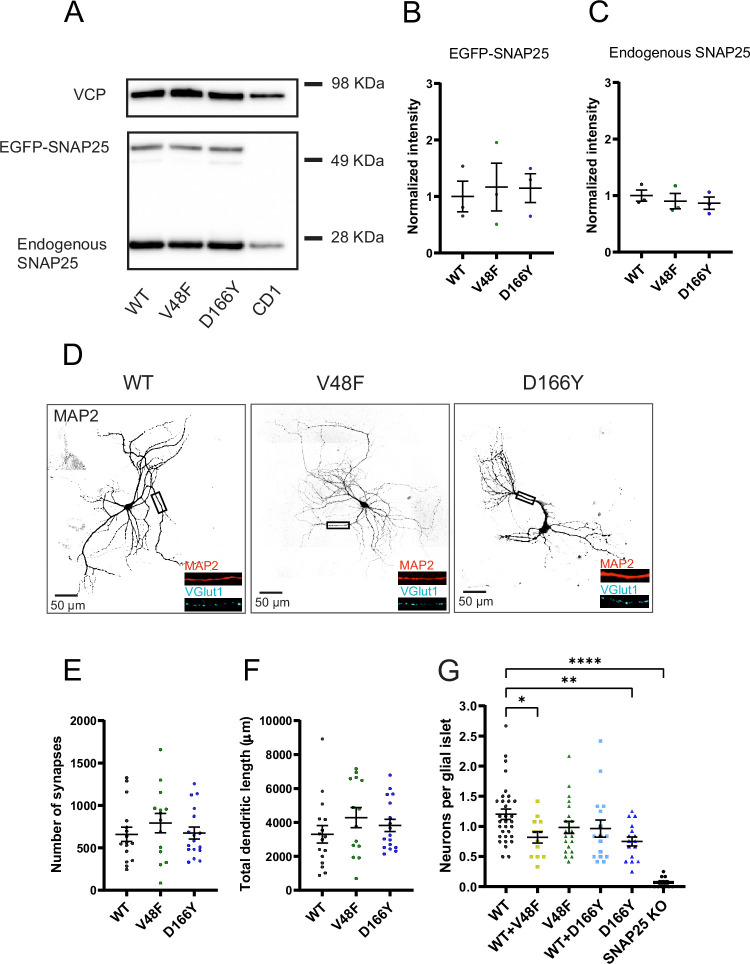
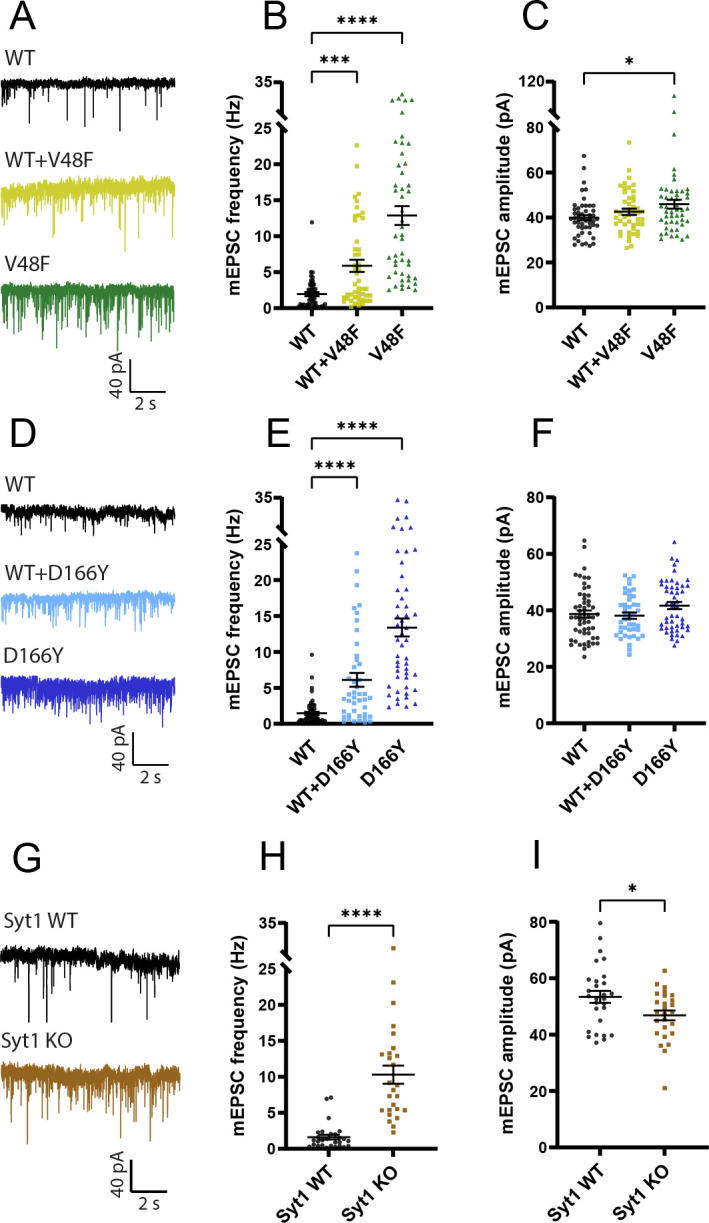
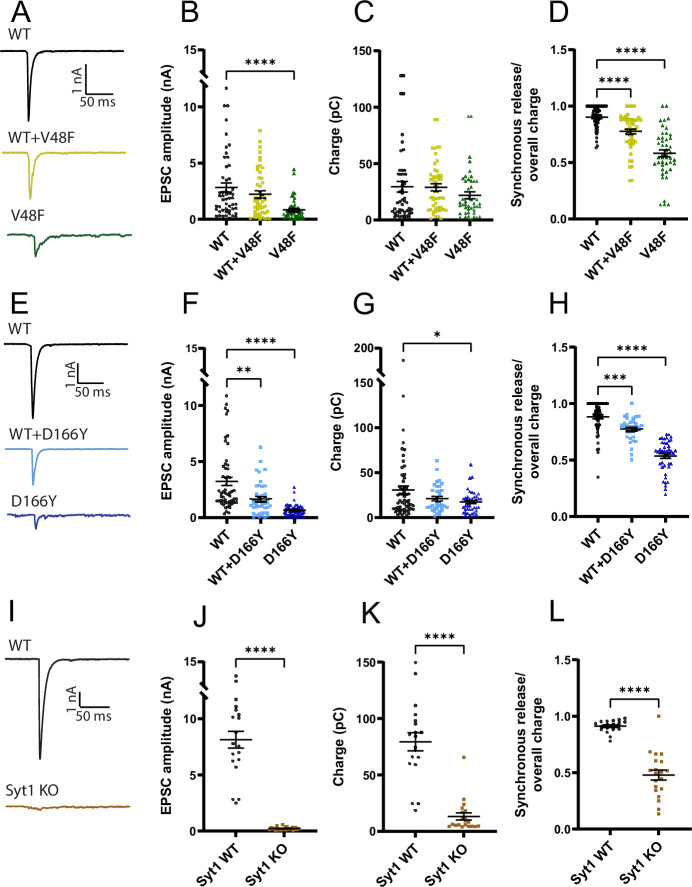
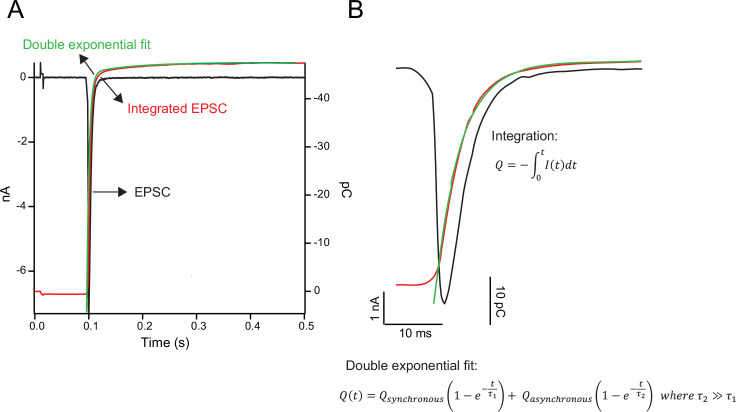
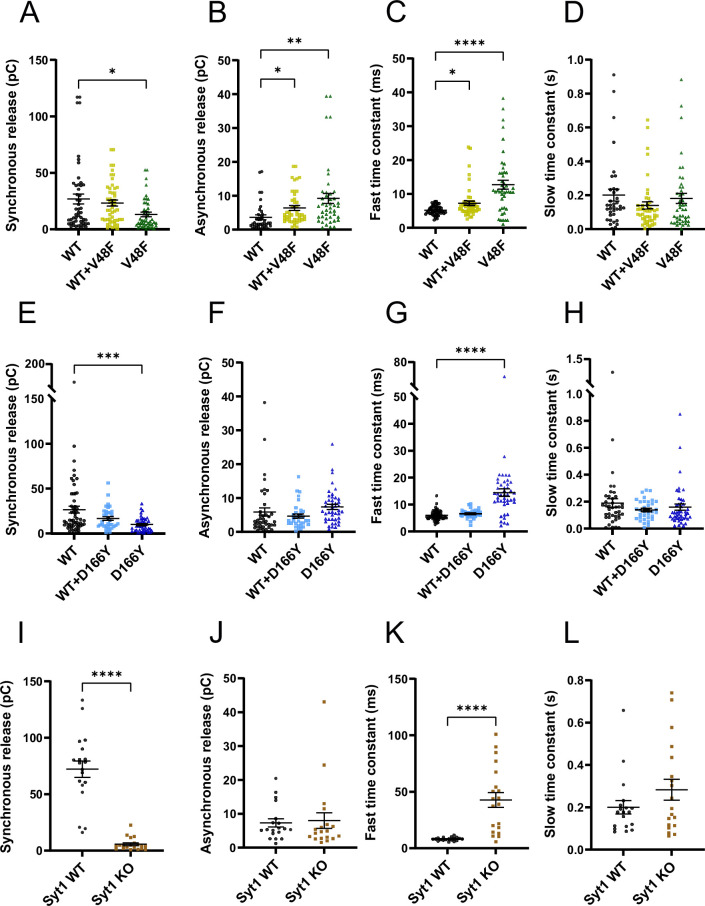
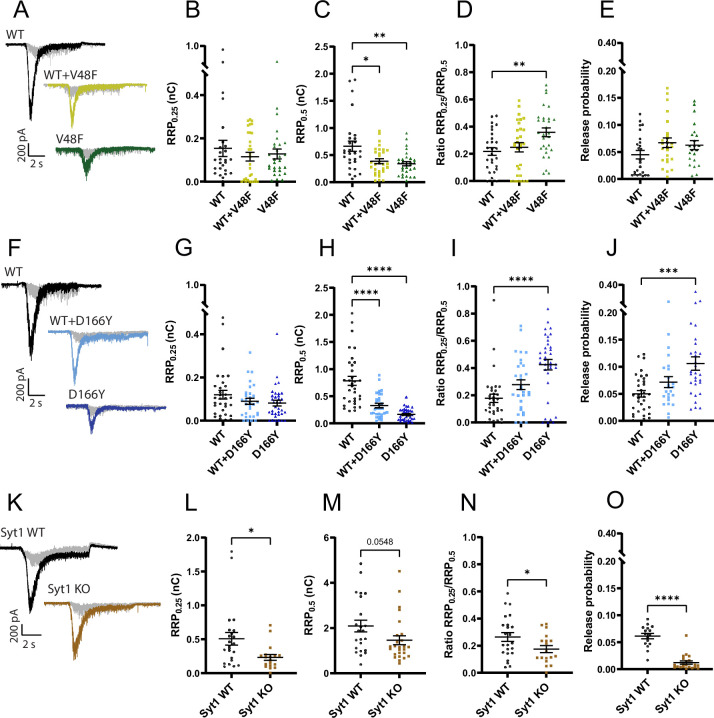
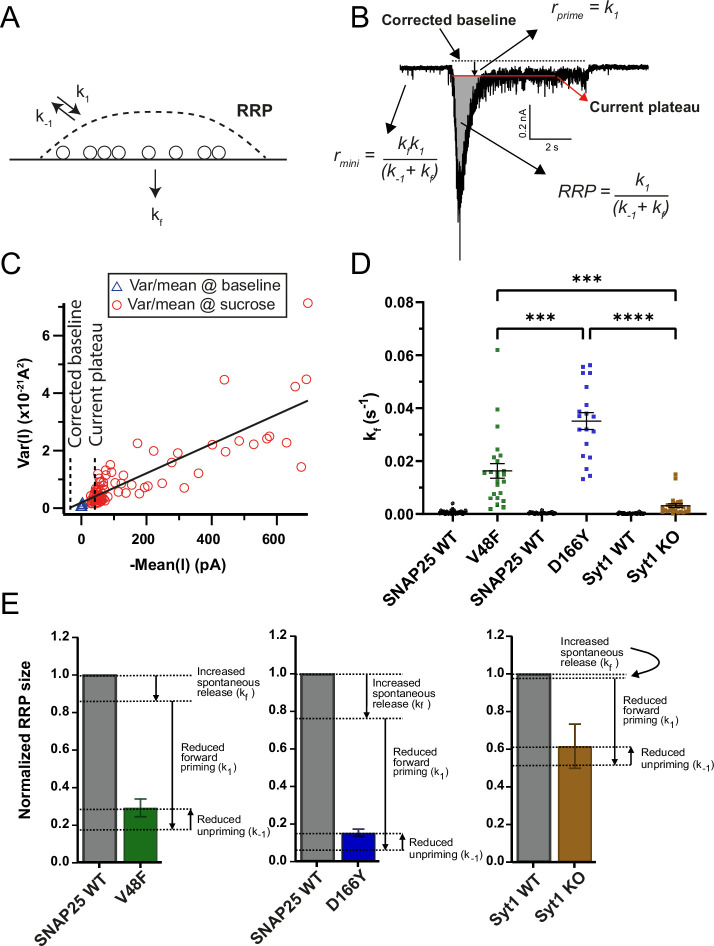
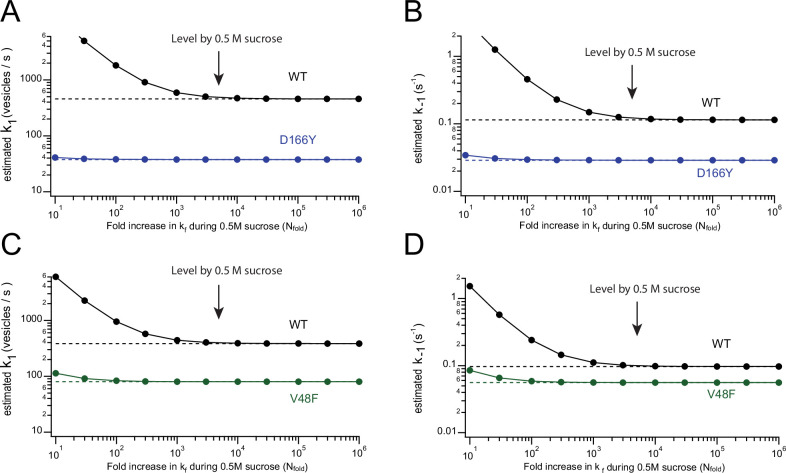
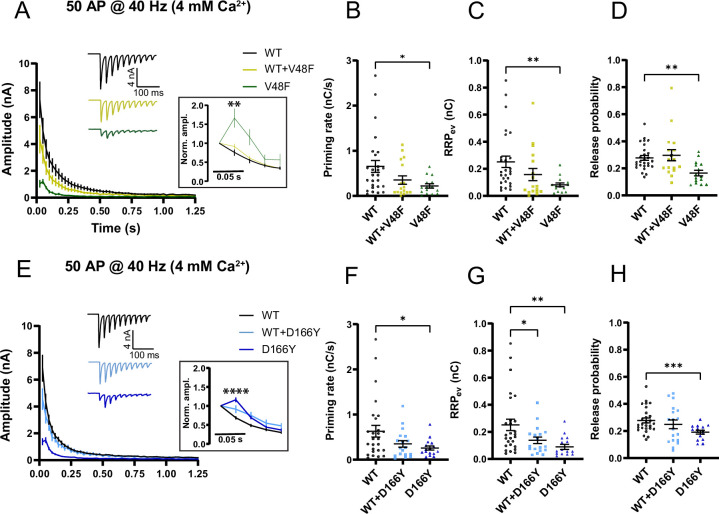
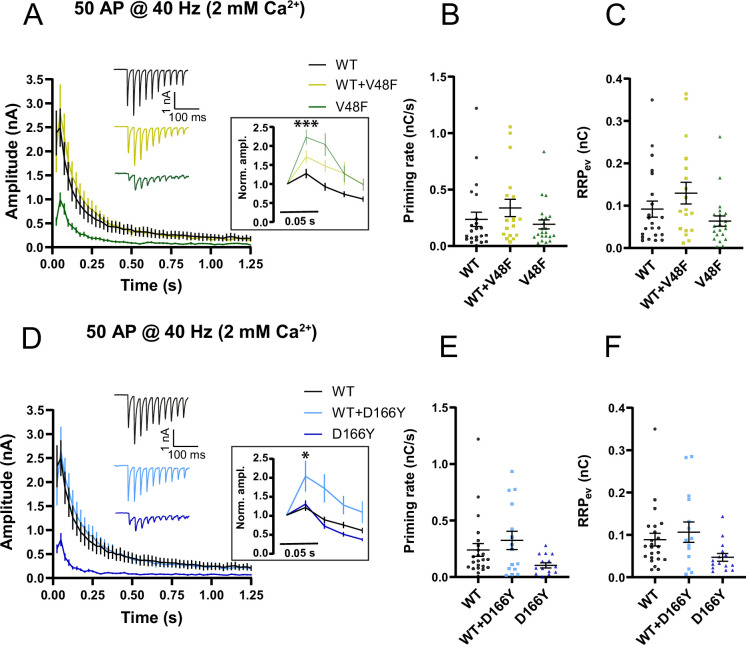
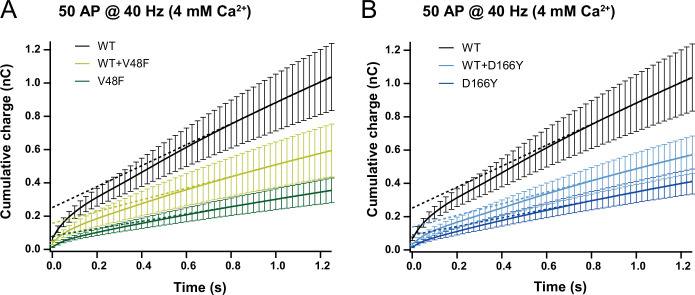
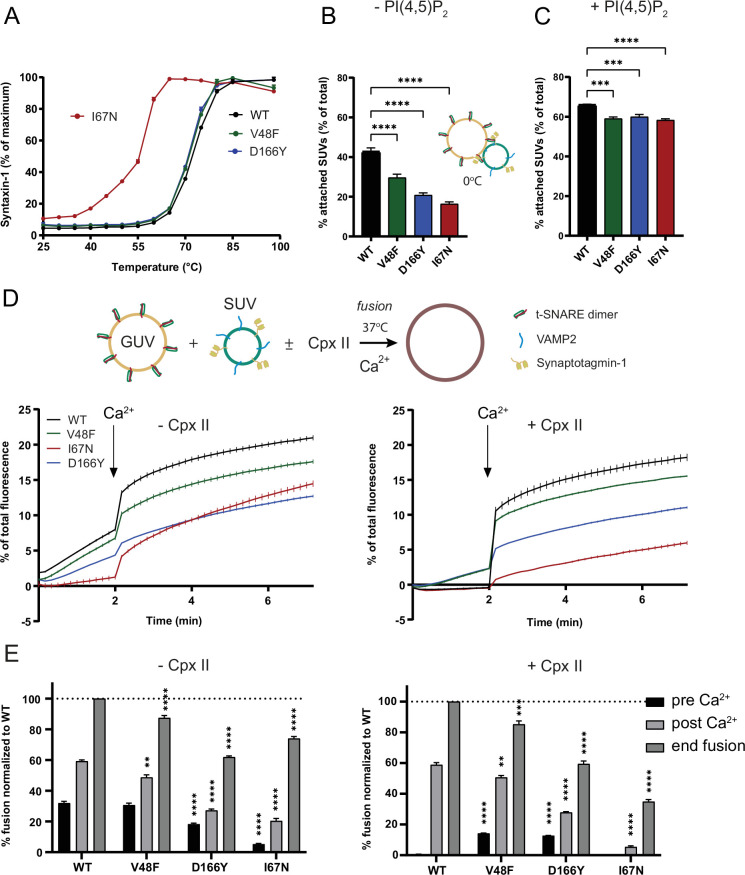
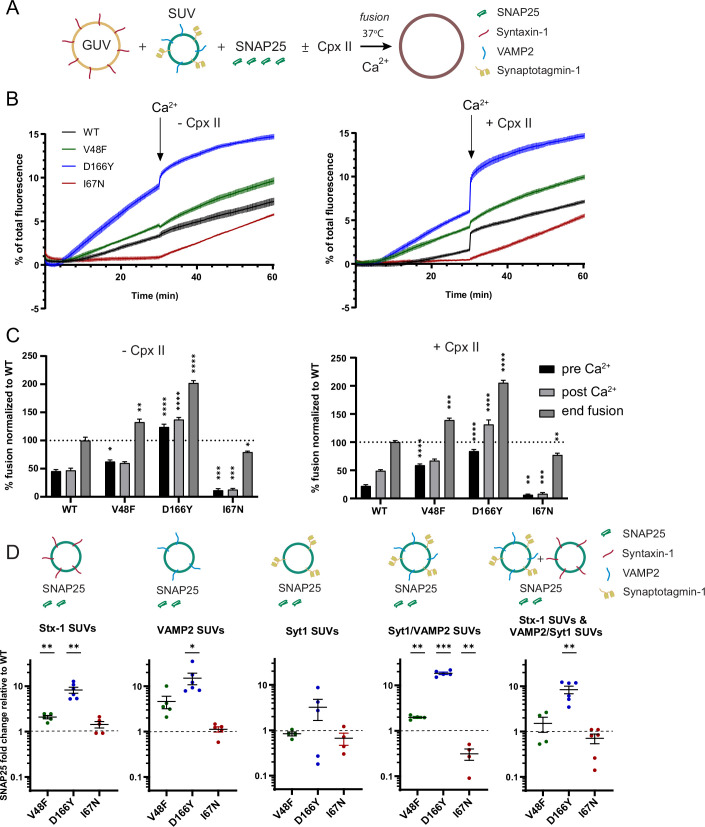
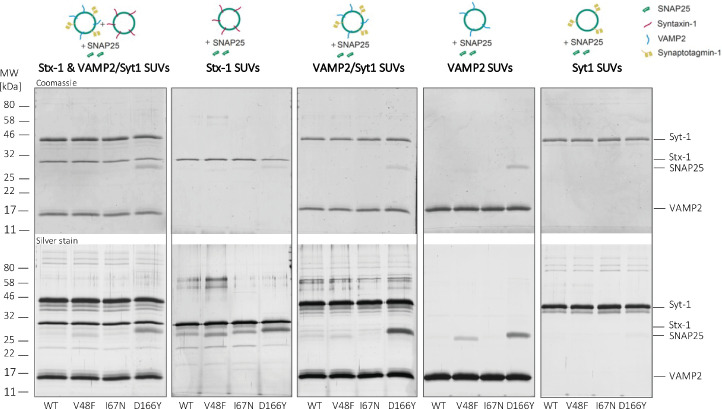
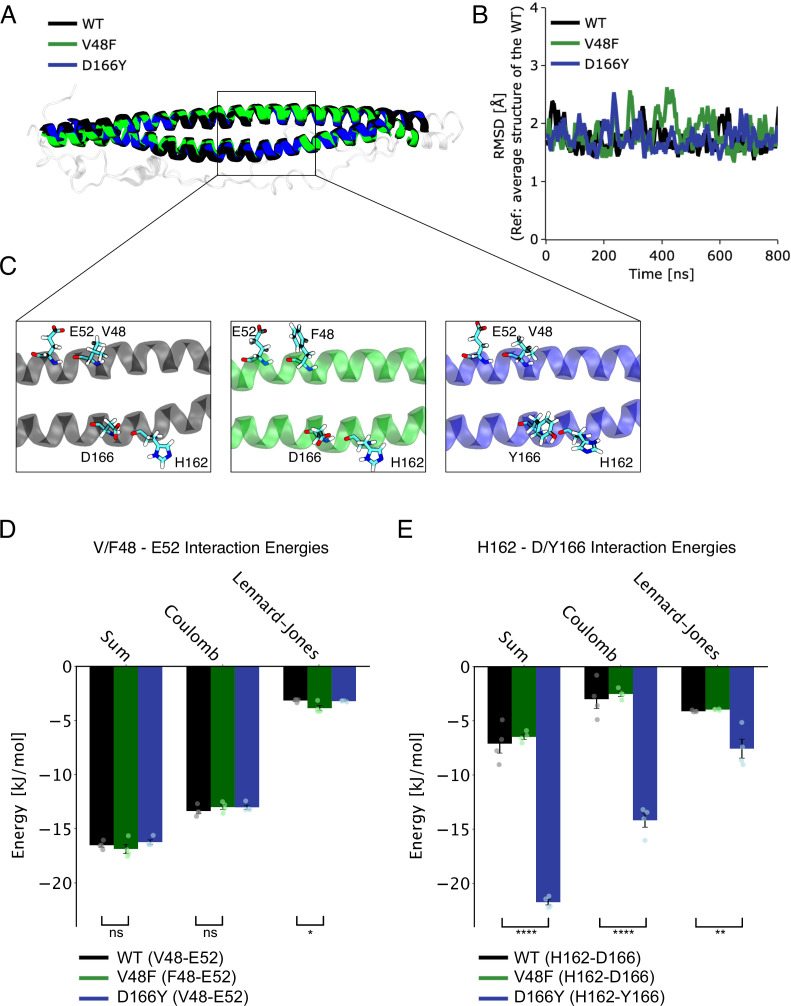
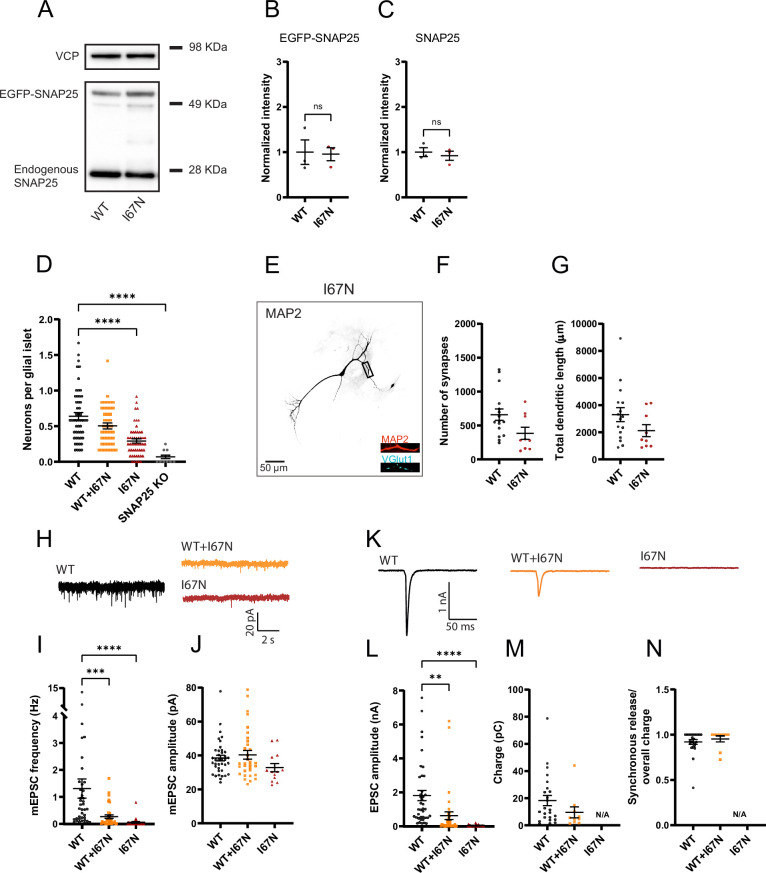
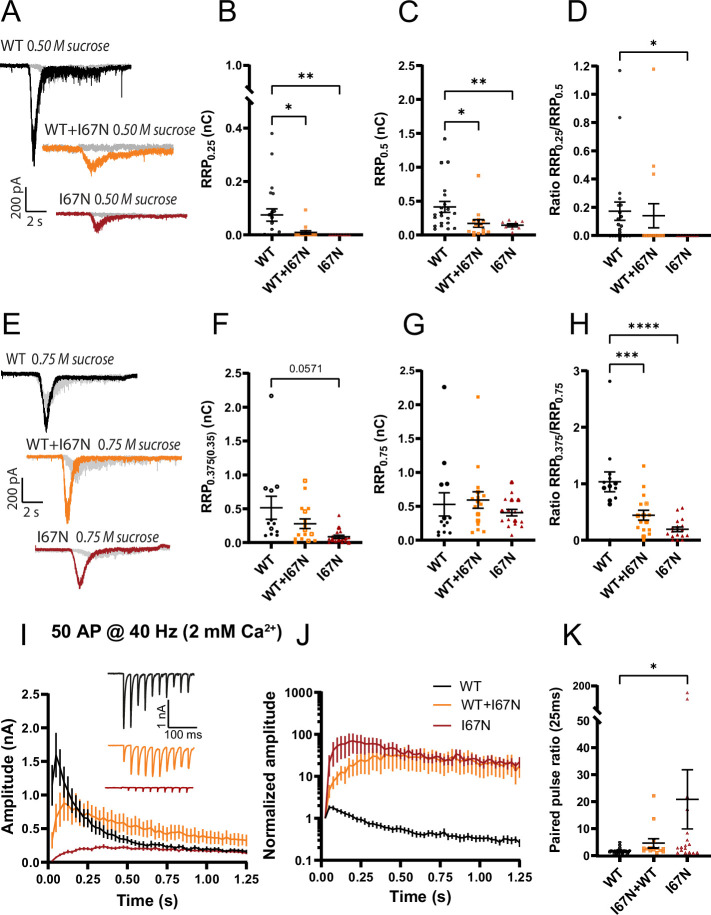
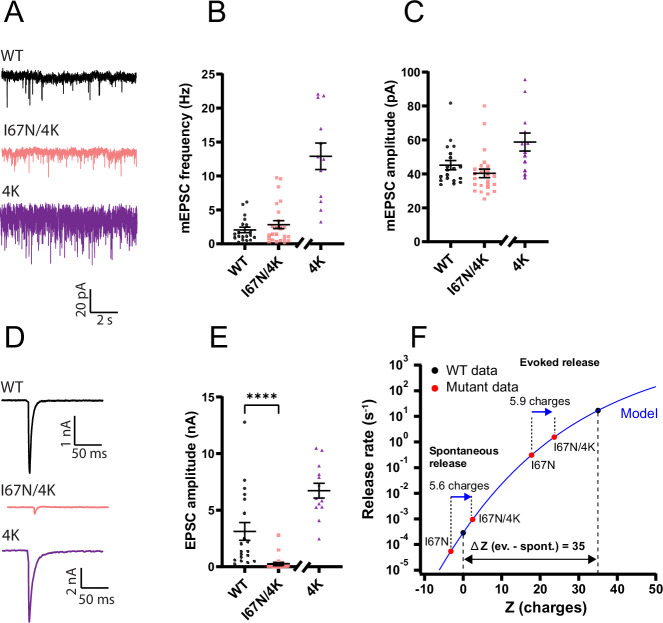
SNAP25 is one of three neuronal SNAREs driving synaptic vesicle exocytosis. We studied three mutations in SNAP25 that cause epileptic encephalopathy: V48F, and D166Y in the synaptotagmin-1 (Syt1)-binding interface, and I67N, which destabilizes the SNARE complex. All three mutations reduced Syt1-dependent vesicle docking to SNARE-carrying liposomes and Ca-stimulated membrane fusion in vitro and when expressed in mouse hippocampal neurons. The V48F and D166Y mutants (with potency D166Y > V48F) led to reduced readily releasable pool (RRP) size, due to increased spontaneous (miniature Excitatory Postsynaptic Current, mEPSC) release and decreased priming rates. These mutations lowered the energy barrier for fusion and increased the release probability, which are gain-of-function features not found in knockout (KO) neurons; normalized mEPSC release rates were higher (potency D166Y > V48F) than in the KO. These mutations (potency D166Y > V48F) increased spontaneous association to partner SNAREs, resulting in unregulated membrane fusion. In contrast, the I67N mutant decreased mEPSC frequency and evoked EPSC amplitudes due to an increase in the height of the energy barrier for fusion, whereas the RRP size was unaffected. This could be partly compensated by positive charges lowering the energy barrier. Overall, pathogenic mutations in SNAP25 cause complex changes in the energy landscape for priming and fusion.
SNAP25 是三种驱动突触小泡胞吐的神经元 SNARE 之一。我们研究了导致癫痫性脑病的 SNAP25 的三种突变:位于突触融合蛋白 1(Syt1)结合界面的 V48F 和 D166Y,以及使 SNARE 复合物不稳定的 I67N。这三种突变都减少了 Syt1 依赖性囊泡停泊在携带 SNARE 的脂质体上的能力,并且在体外以及在表达于小鼠海马神经元时减少了 Ca2+刺激的膜融合。V48F 和 D166Y 突变体(D166Y 的效力>V48F)导致易释放池(RRP)的大小减小,这是由于自发释放(微小兴奋性突触后电流,mEPSC)增加和引发率降低所致。这些突变降低了融合的能垒并增加了释放概率,这些都是在 knockout(KO)神经元中未发现的功能获得特征;正常化的 mEPSC 释放率更高(D166Y 的效力>V48F)比 KO 神经元。这些突变(D166Y 的效力>V48F)增加了与伴侣 SNARE 的自发结合,导致不受调节的膜融合。相比之下,I67N 突变体由于融合能垒的高度增加而降低了 mEPSC 的频率和诱发的 EPSC 幅度,而 RRP 的大小不受影响。这部分可以通过降低能垒的正电荷来部分补偿。总体而言,SNAP25 的致病突变导致引发和融合的能量景观发生复杂变化。