Univ. Grenoble Alpes, CNRS, CEA, Institut de Biologie Structurale, Grenoble, France.
European Molecular Biology Laboratory, Grenoble, France.
Elife. 2024 Apr 19;13:RP93759. doi: 10.7554/eLife.93759.
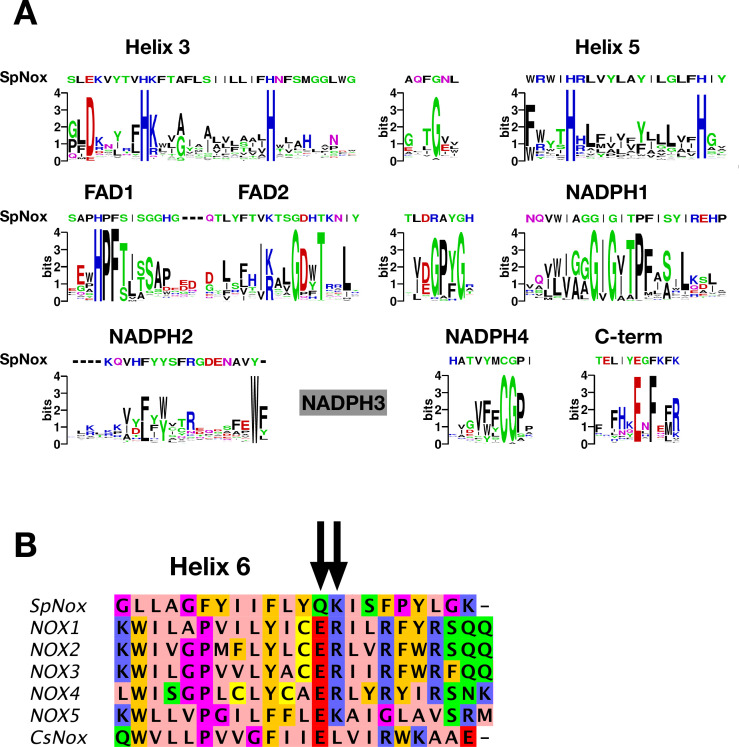
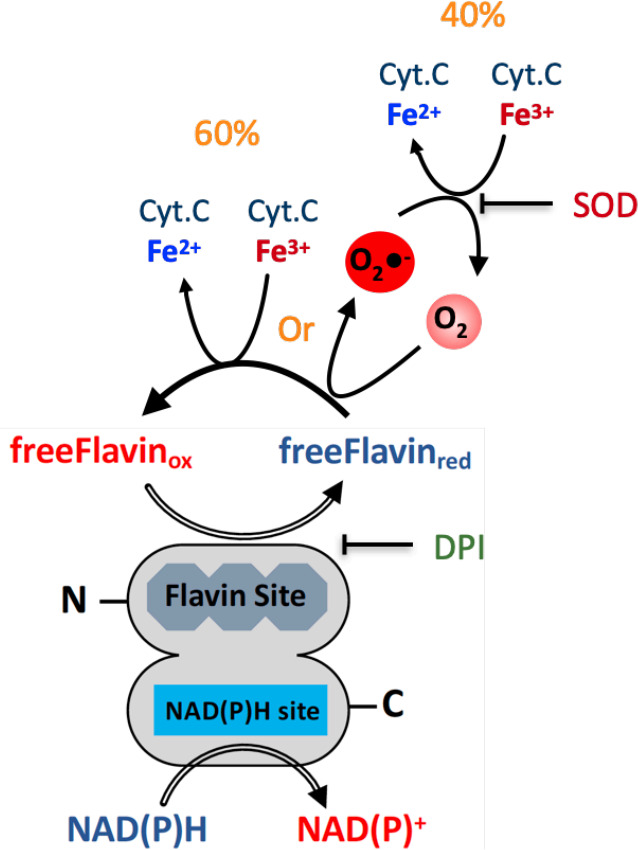
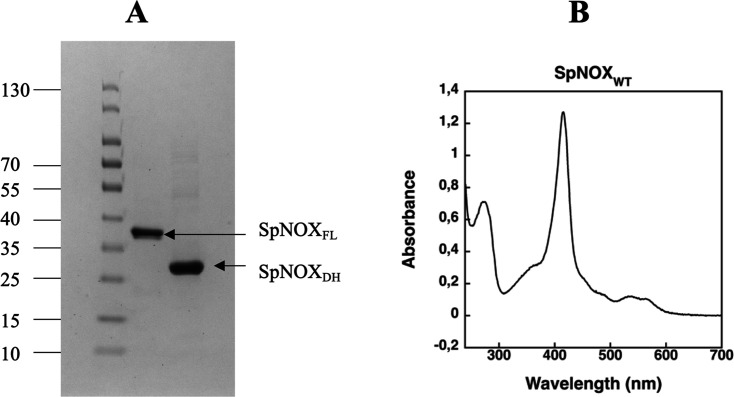
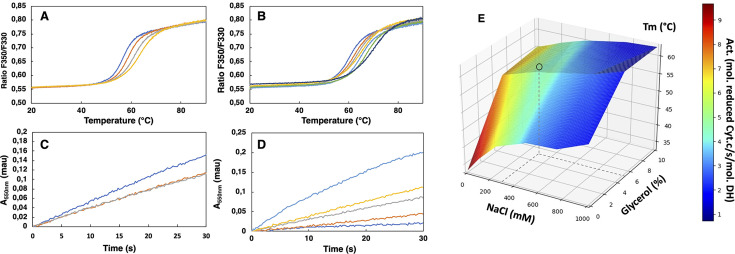
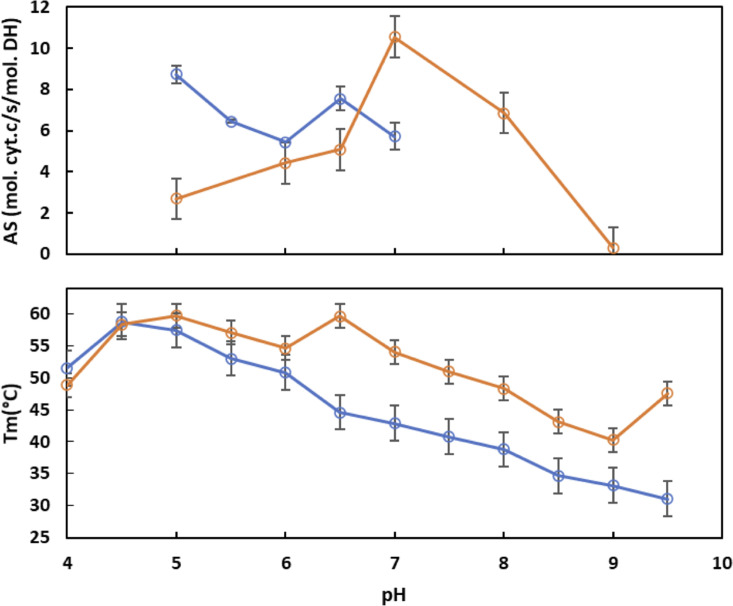
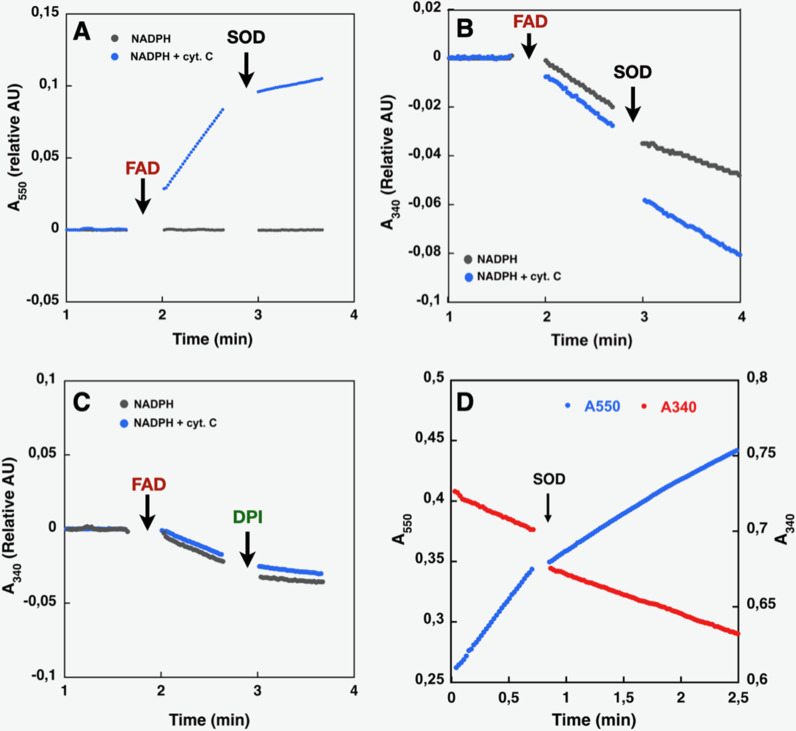
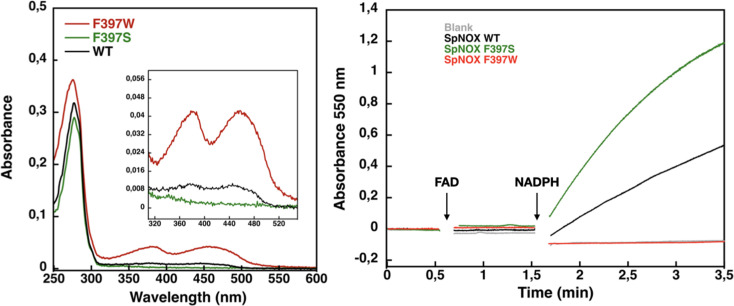
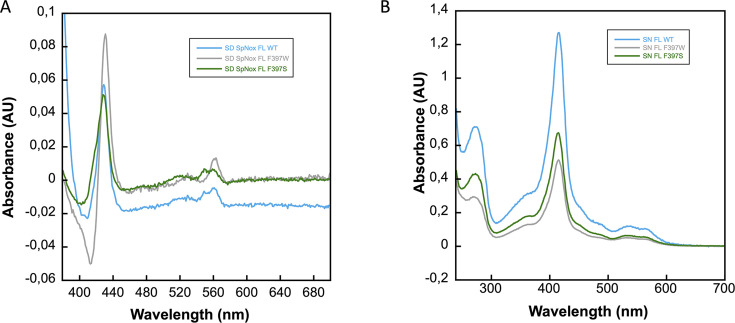
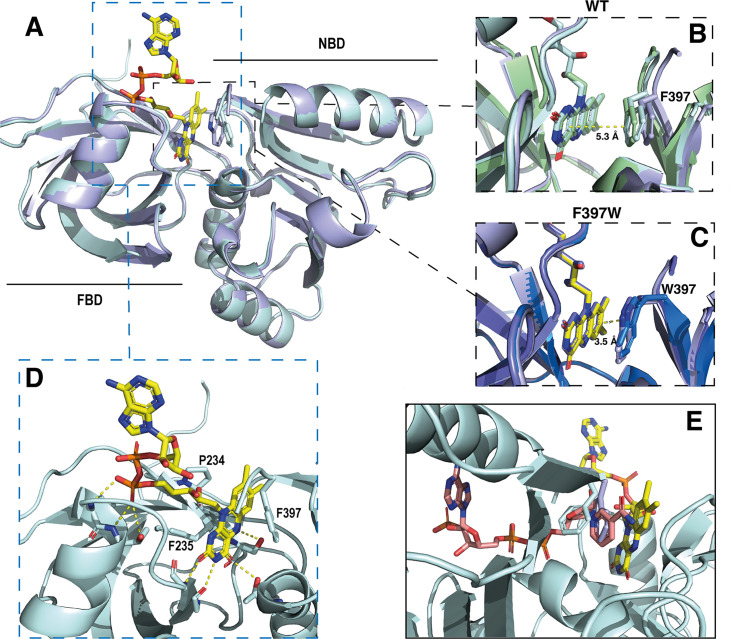
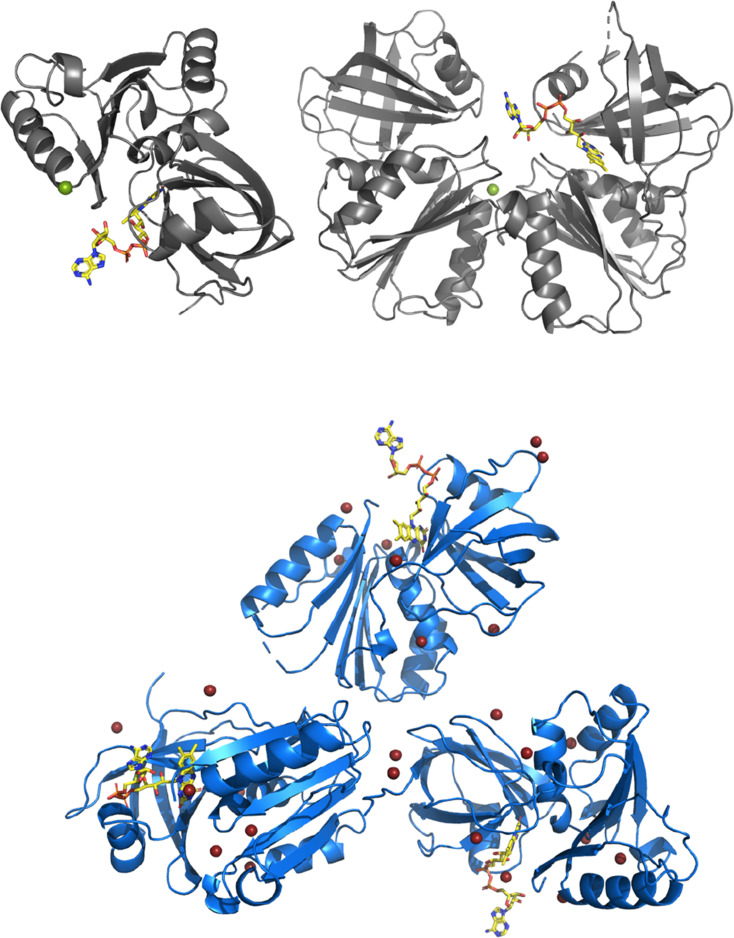
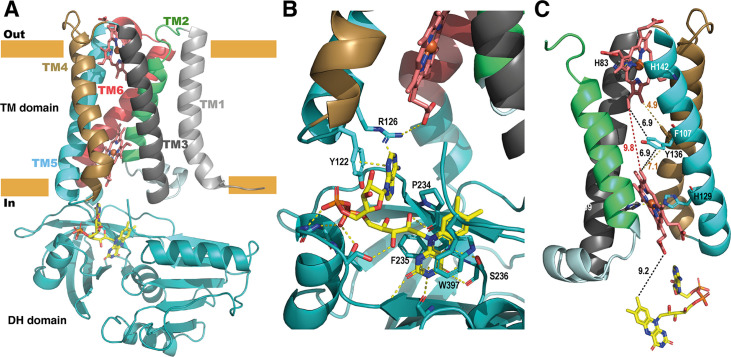
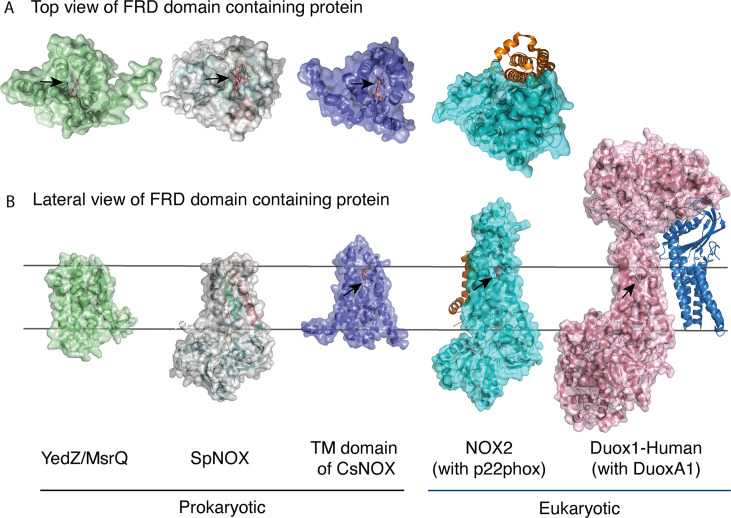
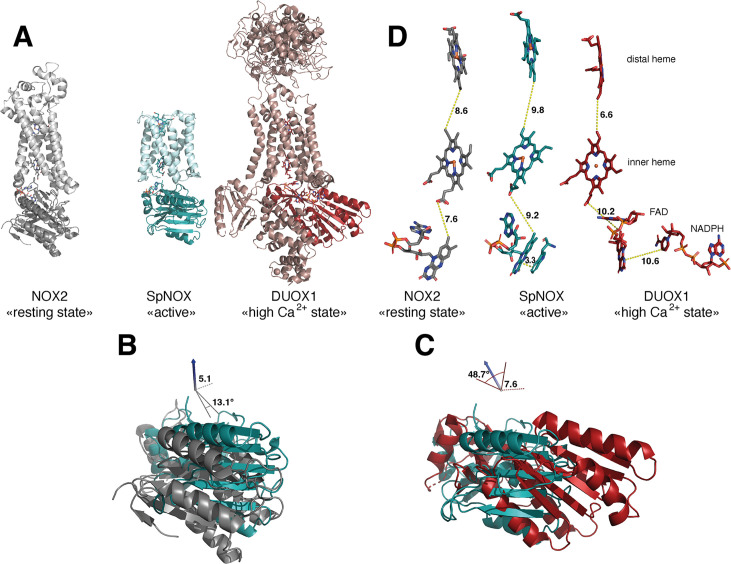
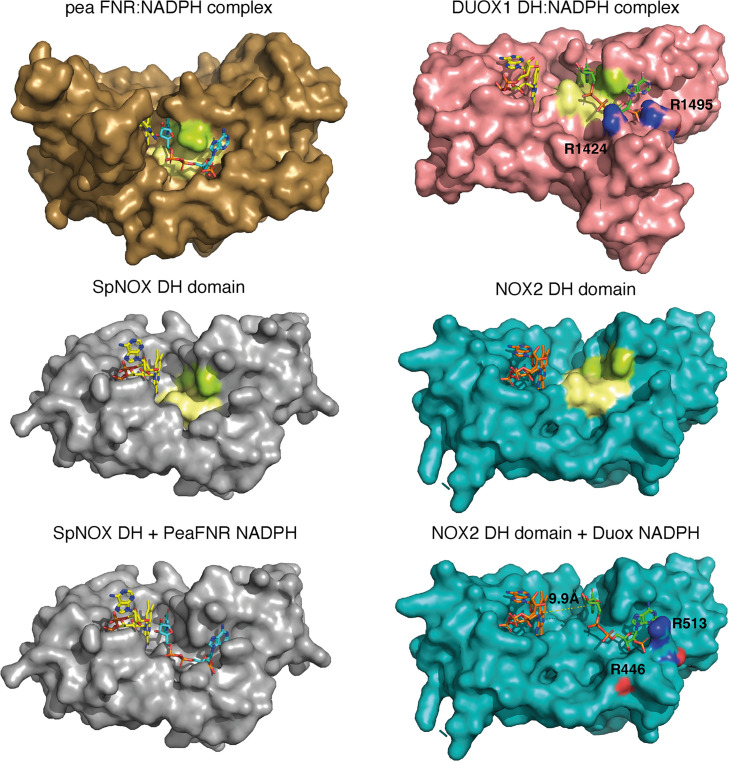
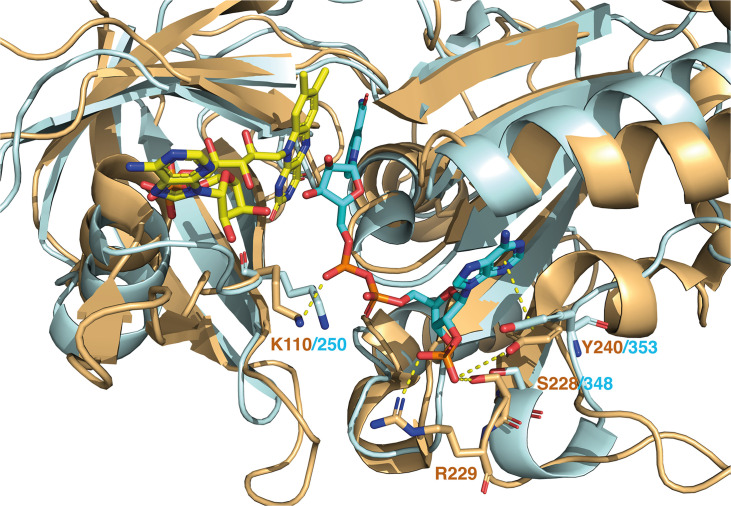
NADPH oxidases (NOX) are transmembrane proteins, widely spread in eukaryotes and prokaryotes, that produce reactive oxygen species (ROS). Eukaryotes use the ROS products for innate immune defense and signaling in critical (patho)physiological processes. Despite the recent structures of human NOX isoforms, the activation of electron transfer remains incompletely understood. SpNOX, a homolog from , can serves as a robust model for exploring electron transfers in the NOX family thanks to its constitutive activity. Crystal structures of SpNOX full-length and dehydrogenase (DH) domain constructs are revealed here. The isolated DH domain acts as a flavin reductase, and both constructs use either NADPH or NADH as substrate. Our findings suggest that hydride transfer from NAD(P)H to FAD is the rate-limiting step in electron transfer. We identify significance of F397 in nicotinamide access to flavin isoalloxazine and confirm flavin binding contributions from both DH and Transmembrane (TM) domains. Comparison with related enzymes suggests that distal access to heme may influence the final electron acceptor, while the relative position of DH and TM does not necessarily correlate with activity, contrary to previous suggestions. It rather suggests requirement of an internal rearrangement, within the DH domain, to switch from a resting to an active state. Thus, SpNOX appears to be a good model of active NOX2, which allows us to propose an explanation for NOX2's requirement for activation.
NADPH 氧化酶(NOX)是一种跨膜蛋白,广泛存在于真核生物和原核生物中,可产生活性氧(ROS)。真核生物利用 ROS 产物进行先天免疫防御和关键(病理)生理过程中的信号转导。尽管最近已经解析了人类 NOX 同工型的结构,但电子传递的激活仍未完全了解。来自 的 SpNOX 同源物可以作为探索 NOX 家族中电子传递的强大模型,因为其具有组成型活性。本文揭示了全长 SpNOX 和脱氢酶(DH)结构域构建体的晶体结构。分离的 DH 结构域充当黄素还原酶,并且这两种构建体都可以使用 NADPH 或 NADH 作为底物。我们的研究结果表明,从 NAD(P)H 到 FAD 的氢化物转移是电子传递的限速步骤。我们确定了 F397 在烟酰胺进入黄素异咯嗪中的重要性,并证实了 DH 和跨膜(TM)结构域都对黄素结合有贡献。与相关酶的比较表明,远端进入血红素可能影响最终电子受体,而 DH 和 TM 的相对位置不一定与活性相关,这与之前的建议相反。它表明需要在 DH 结构域内进行内部重排,以从静止状态切换到激活状态。因此,SpNOX 似乎是活性 NOX2 的良好模型,这使我们能够对 NOX2 的激活要求提出解释。