Department of Internal Medicine and Pediatrics, Ghent University, Ghent, Belgium.
VIB Center for Inflammation Research, Ghent, Belgium.
Front Immunol. 2024 Apr 12;15:1373745. doi: 10.3389/fimmu.2024.1373745. eCollection 2024.
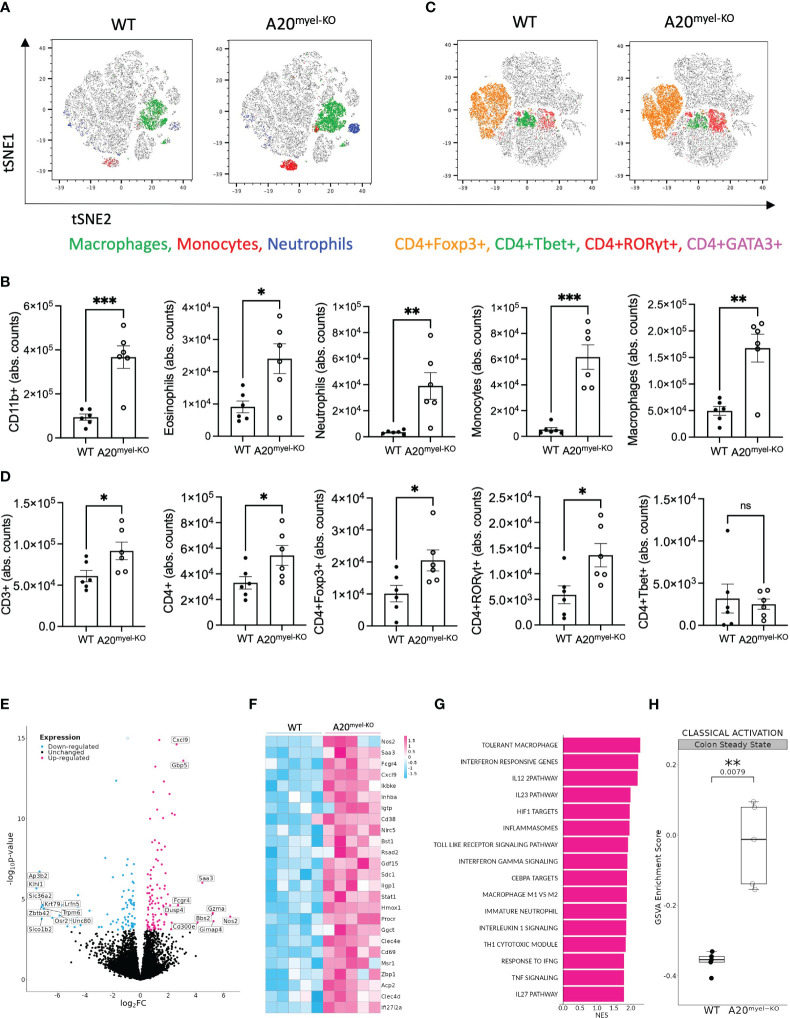
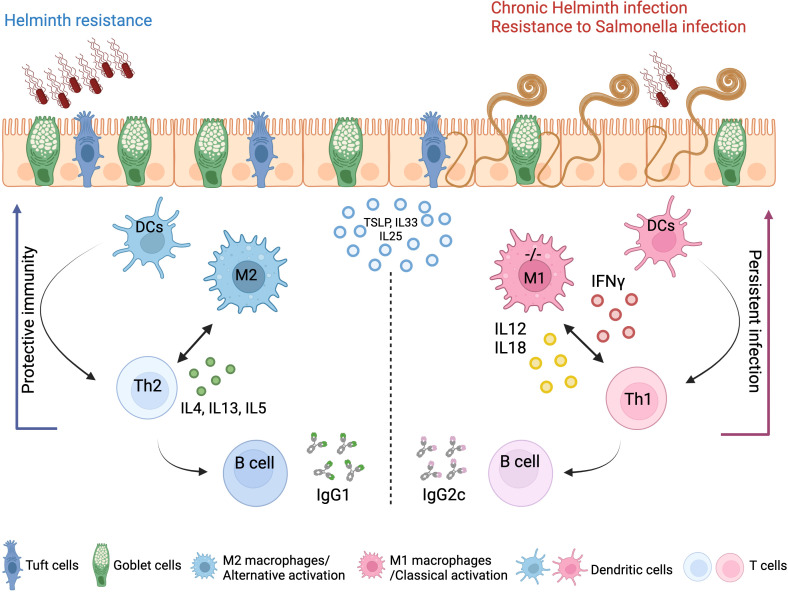
Protective immunity against intestinal helminths requires induction of robust type-2 immunity orchestrated by various cellular and soluble effectors which promote goblet cell hyperplasia, mucus production, epithelial proliferation, and smooth muscle contractions to expel worms and re-establish immune homeostasis. Conversely, defects in type-2 immunity result in ineffective helminth clearance, persistent infection, and inflammation. Macrophages are highly plastic cells that acquire an alternatively activated state during helminth infection, but they were previously shown to be dispensable for resistance to infection.
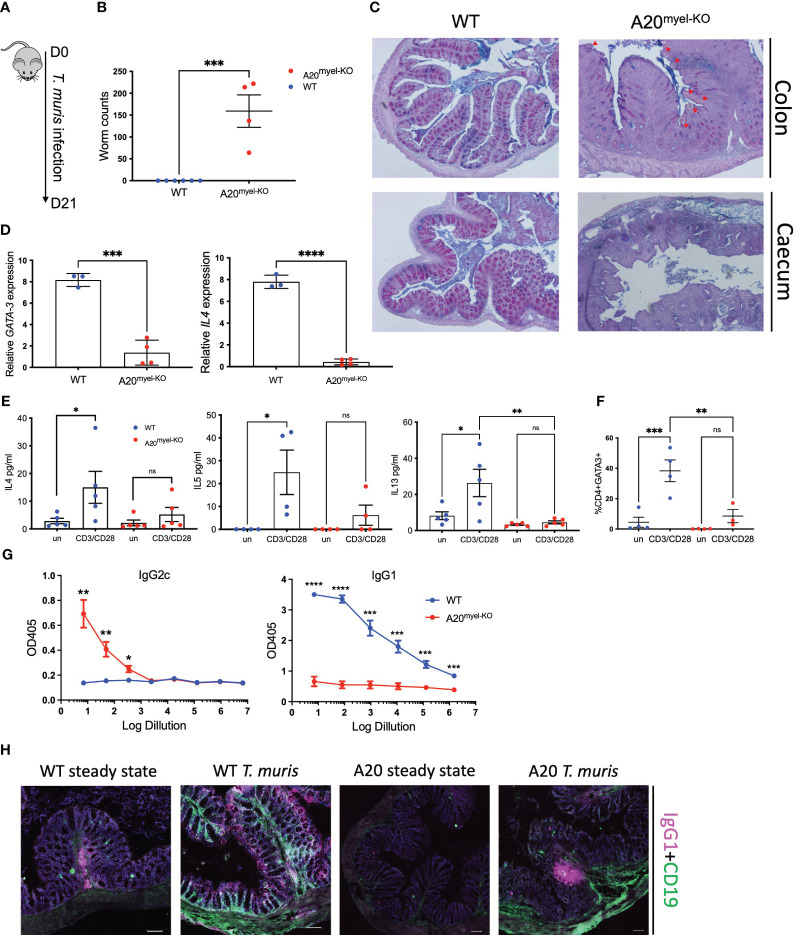
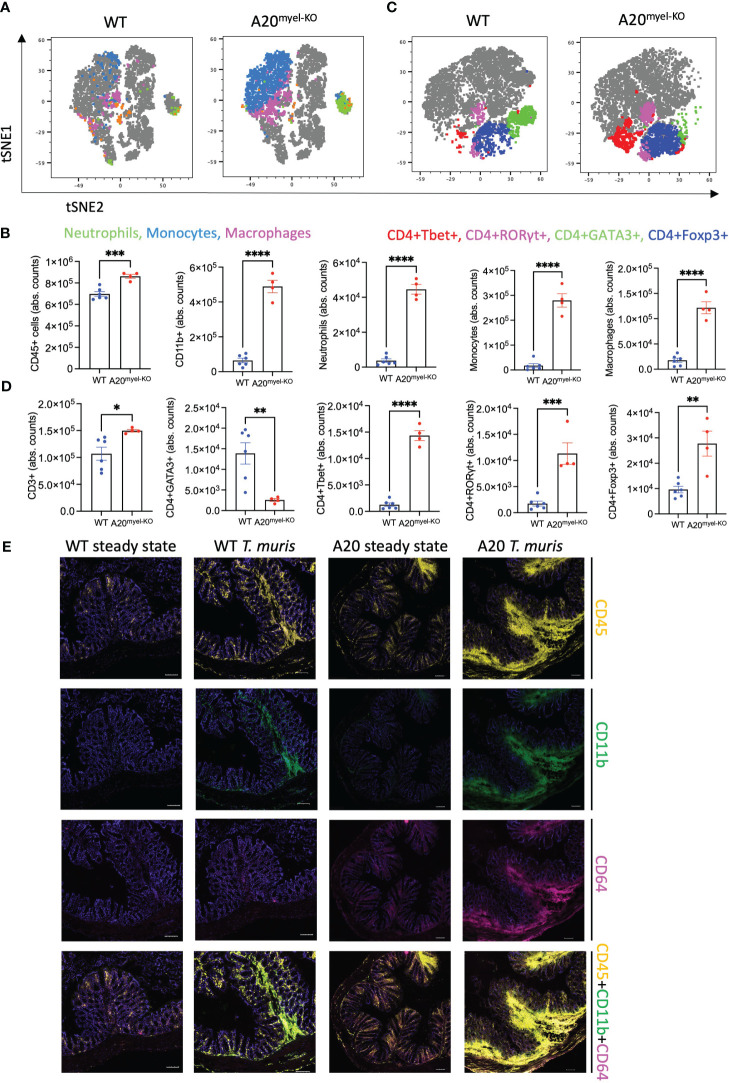
We use the in vivo mouse model A20myel-KO, characterized by the deletion of the potent anti-inflammatory factor A20 (TNFAIP3) specifically in the myeloid cells, the excessive type-1 cytokine production, and the development of spontaneous arthritis. We infect A20 mice with the gastrointestinal helminth and we analyzed the innate and adaptive responses. We performed RNA sequencing on sorted myeloid cells to investigate the role of A20 on macrophage polarization and type-2 immunity. Moreover, we assess in A20 mice the pharmacological inhibition of type-1 cytokine pathways on helminth clearance and the infection with .
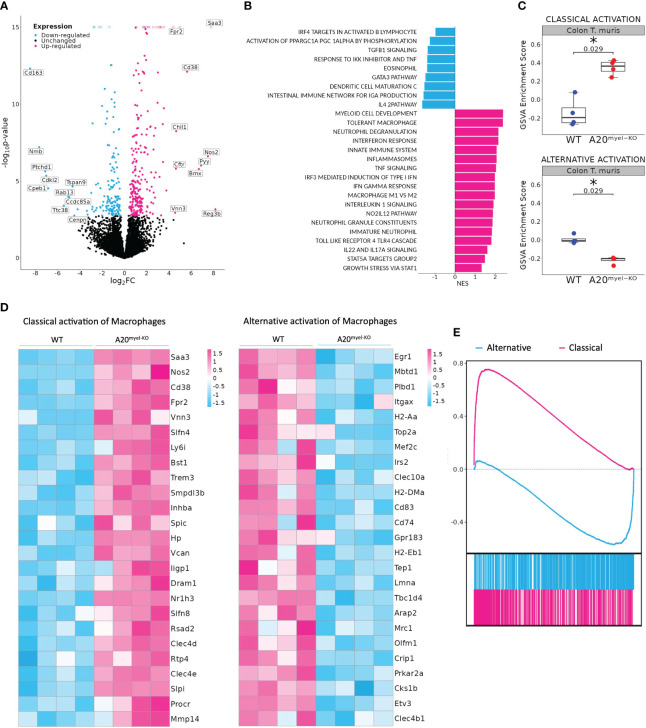
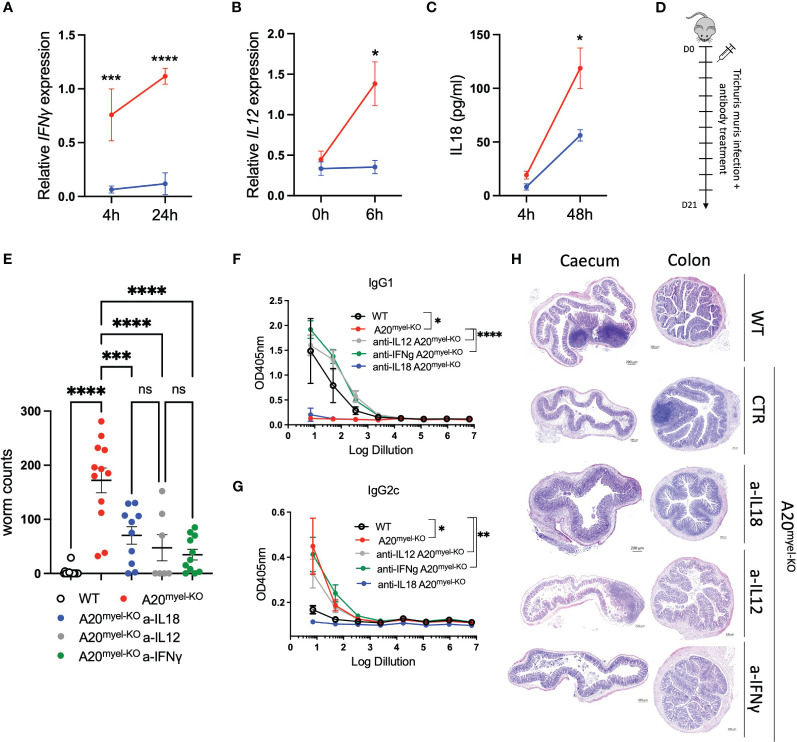
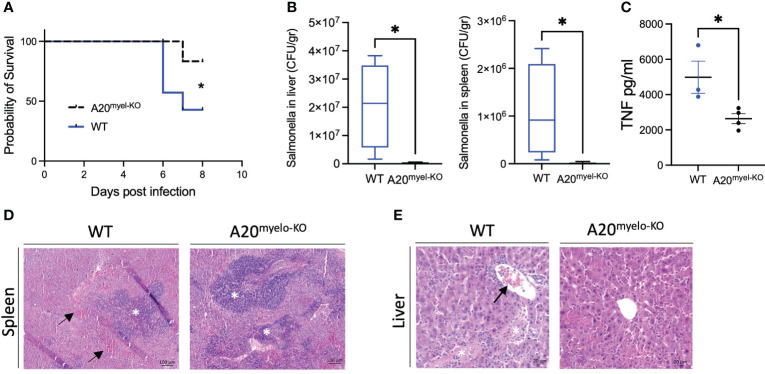
We show that proper macrophage polarization is essential for helminth clearance, and we identify A20 as an essential myeloid factor for the induction of type-2 immune responses against . A20 mice are characterized by persistent infection and intestinal inflammation. Myeloid A20 deficiency induces strong classical macrophage polarization which impedes anti-helminth type-2 immune activation; however, it promotes detrimental Th1/Th17 responses. Antibody-mediated neutralization of the type-1 cytokines IFN-γ, IL-18, and IL-12 prevents myeloid-orchestrated Th1 polarization and re-establishes type-2-mediated protective immunity against in A20 mice. In contrast, the strong Th1-biased immunity in A20 mice offers protection against infection.
We hereby identify A20 as a critical myeloid factor for correct macrophage polarization and appropriate adaptive mucosal immunity in response to helminth and enteric bacterial infection.
针对肠道寄生虫的保护性免疫需要由各种细胞和可溶性效应物诱导产生强大的 2 型免疫,促进杯状细胞增生、黏液产生、上皮细胞增殖和平滑肌收缩,以驱除蠕虫并重新建立免疫稳态。相反,2 型免疫缺陷会导致蠕虫清除无效、持续感染和炎症。巨噬细胞是高度可塑性的细胞,在寄生虫感染时会获得一种替代性激活状态,但之前的研究表明它们对于抵抗感染是可有可无的。
我们使用体内小鼠模型 A20myel-KO,其特征是在髓样细胞中特异性缺失强效抗炎因子 A20(TNFAIP3),导致过度的 1 型细胞因子产生和自发性关节炎的发展。我们用胃肠道寄生虫 感染 A20 小鼠,并分析了先天和适应性反应。我们对分选的髓样细胞进行 RNA 测序,以研究 A20 在巨噬细胞极化和 2 型免疫中的作用。此外,我们评估了在 A20 小鼠中,通过抑制 1 型细胞因子通路,对寄生虫清除和 感染的影响。
我们表明,适当的巨噬细胞极化对于寄生虫清除至关重要,并且我们确定 A20 是诱导针对 寄生虫的 2 型免疫反应的必需髓样因子。A20 小鼠的特征是持续的 感染和肠道炎症。髓样 A20 缺陷诱导强烈的经典巨噬细胞极化,阻碍了抗寄生虫的 2 型免疫激活;然而,它促进了有害的 Th1/Th17 反应。中和 1 型细胞因子 IFN-γ、IL-18 和 IL-12 的抗体可防止髓样细胞协调的 Th1 极化,并在 A20 小鼠中重新建立针对 感染的 2 型介导的保护性免疫。相比之下,A20 小鼠强烈的 Th1 偏向性免疫提供了对 感染的保护。
我们确定 A20 是一种关键的髓样因子,可促进针对寄生虫和肠道细菌感染的正确巨噬细胞极化和适当的适应性黏膜免疫。