Department of Plant Ecology, Faculty of Biology, University of Gdansk, 59 Wita Stwosza St., Gdańsk, PL, 80-308, Poland.
Department of Plant Experimental Biology and Biotechnology, Faculty of Biology, University of Gdansk, 59 Wita Stwosza St., Gdańsk, 80-308, Poland.
BMC Plant Biol. 2024 May 23;24(1):449. doi: 10.1186/s12870-024-05155-9.
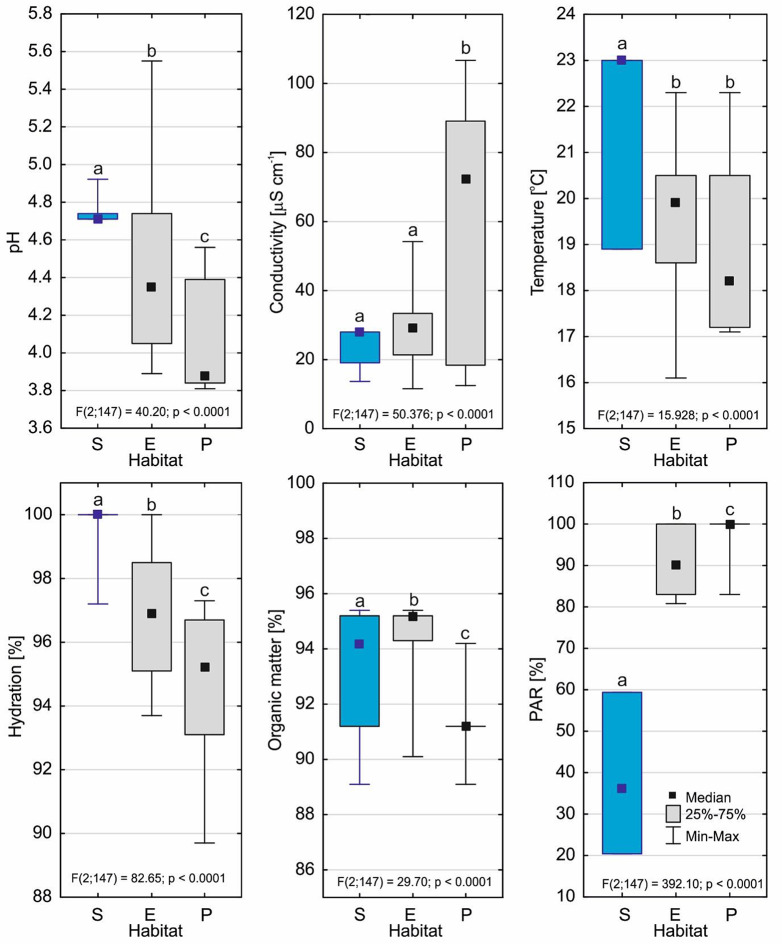
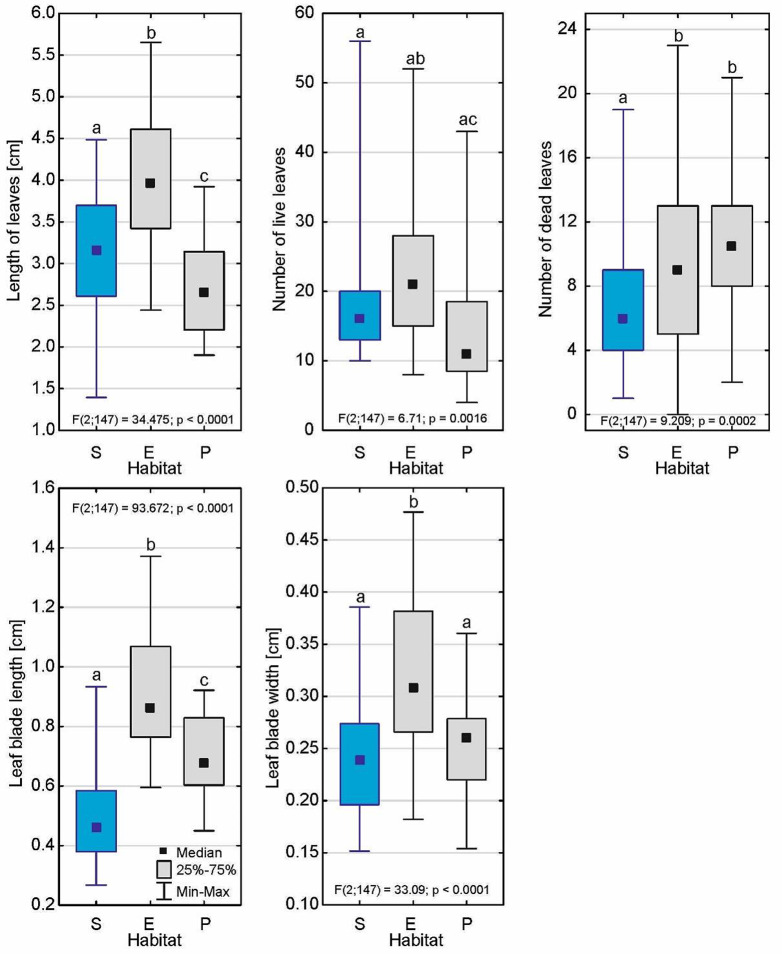
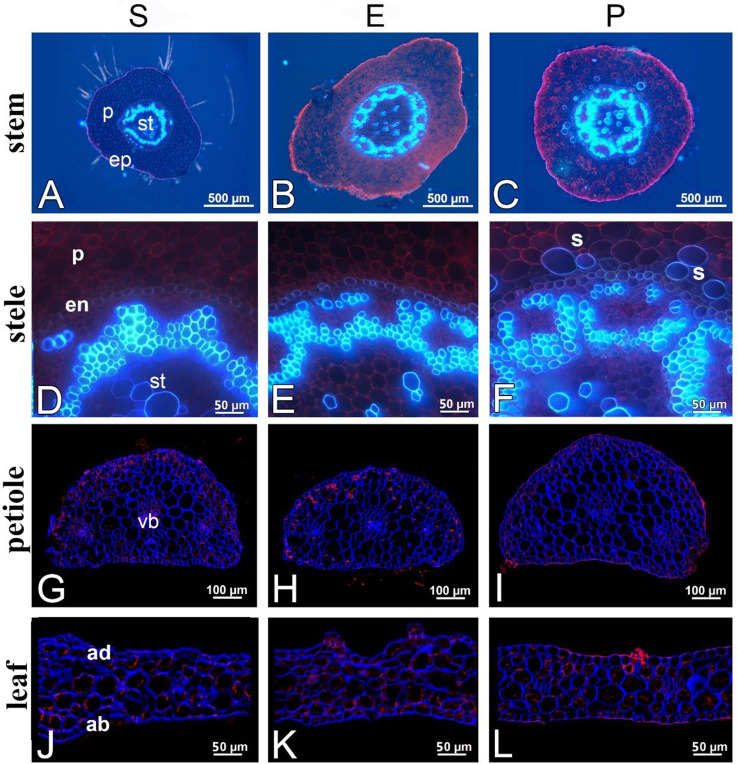
Drosera intermedia grows in acidic bogs in parts of valleys that are flooded in winter, and that often dry out in summer. It is also described as the sundew of the most heavily hydrated habitats in peatlands, and it is often found in water and even underwater. This sundew is the only one that can tolerate long periods of submersion, and more importantly produces a typical submerged form that can live in such conditions for many years. Submerged habitats are occupied by D. intermedia relatively frequently. The aim of the study was to determine the environmental conditions and architecture of individuals in the submerged form of D. intermedia. The features of the morphological and anatomical structure and chlorophyll a fluorescence of this form that were measured were compared with analogous ones in individuals that occurred in emerged and peatland habitats. The submerged form occurred to a depth of 20 cm. Compared to the other forms, its habitat had the highest pH (4.71-4.92; Me = 4.71), the highest temperature and substrate hydration, and above all, the lowest photosynthetically active radiation (PAR; 20.4-59.4%). This form differed from the other forms in almost all of the features of the plant's architecture. It is particularly noteworthy that it had the largest main axis height among all of the forms, which exceeded 18 cm. The number of living leaves in a rosette was notable (18.1 ± 8.1), while the number of dead leaves was very low (6.9 ± 3.8). The most significant differences were in the shape of its submerged leaves, in which the length of the leaf blade was the lowest of all of the forms (0.493 ± 0.15 mm; p < 0.001) and usually the widest. The stem cross-sectional area was noticeably smaller in the submerged form than in the other forms, the xylem was less developed and collaterally closed vascular bundles occurred. Our analysis of the parameters of chlorophyll fluorescence in vivo revealed that the maximum quantum yield of the primary photochemistry of photosystem II is the highest for the submerged form (Me = 0.681), the same as the maximum quantum yield of the electron transport (Me φE = 0.183). The efficiency of energy use per one active reaction center of photosystem II (RC) was the lowest in the submerged form (Me = 2.978), same as the fraction of energy trapped by one active RC (Me = 1.976) and the non-photochemical energy dissipation (DI/RC; Me = 0.916). The ET/RC parameter, associated with the efficiency of the energy utilization for electron transport by one RC, in the submerged plant reached the highest value (Me = 0.489). The submerged form of D. intermedia clearly differed from the emerged and peatland forms in its plant architecture. The submerged plants had a thinner leaf blade and less developed xylem than the other forms, however, their stems were much longer. The relatively high photosynthetic efficiency of the submerged forms suggests that most of the trapped energy is utilized to drive photosynthesis with a minimum energy loss, which may be a mechanism to compensate for the relatively small size of the leaf blade.
中肋毛毡苔生长在冬季被洪水淹没的山谷中酸性沼泽地,夏季往往会干涸。它也被描述为泥炭地最湿润生境中的茅膏菜,通常生长在水中,甚至水下。这种茅膏菜是唯一能耐受长时间浸泡的,更重要的是,它能产生一种典型的水下形态,可以在这种条件下生存多年。中肋毛毡苔相对频繁地占据水下栖息地。本研究的目的是确定中肋毛毡苔水下形态的环境条件和个体结构。对该形态的形态和解剖结构以及叶绿素 a 荧光的特征进行了测量,并与在露出水面和泥炭地生境中出现的类似个体进行了比较。水下形态的深度可达 20 厘米。与其他形式相比,其栖息地的 pH 值最高(4.71-4.92;Me = 4.71),温度和基质水化程度最高,最重要的是,光合有效辐射(PAR)最低(20.4-59.4%)。这种形态在植物结构的几乎所有特征上都与其他形态不同。值得注意的是,它的主茎高度是所有形态中最大的,超过 18 厘米。叶丛中活叶的数量(18.1±8.1)是显著的,而死叶的数量非常低(6.9±3.8)。最显著的差异是其水下叶的形状,其中叶片长度是所有形态中最低的(0.493±0.15 毫米;p<0.001),通常也是最宽的。水下形态的茎横截面积明显小于其他形态,木质部发育较差,出现侧向封闭的维管束。我们对体内叶绿素荧光参数的分析表明,光系统 II 原初光化学的最大量子产量对水下形态最高(Me = 0.681),与电子传递的最大量子产量(Me φE = 0.183)相同。每个光系统 II 活性反应中心(RC)利用能量的效率在水下形态中最低(Me = 2.978),与一个活性 RC 捕获的能量分数(Me = 1.976)和非光化学能量耗散(DI/RC;Me = 0.916)相同。与一个 RC 电子传递利用能量的效率相关的 ET/RC 参数在水下植物中达到最高值(Me = 0.489)。中肋毛毡苔的水下形态与露出水面和泥炭地形态在植物结构上明显不同。水下植物的叶片比其他形态更薄,木质部发育程度更低,但茎更长。相对较高的光合作用效率表明,大部分捕获的能量都被用来驱动光合作用,而能量损失最小,这可能是一种补偿叶片相对较小尺寸的机制。