Max Planck Institute for Biology of Ageing, Cologne, Germany.
Cologne Graduate School of Ageing Research, Cologne, Germany.
Nat Cell Biol. 2024 Nov;26(11):1918-1933. doi: 10.1038/s41556-024-01523-7. Epub 2024 Oct 9.
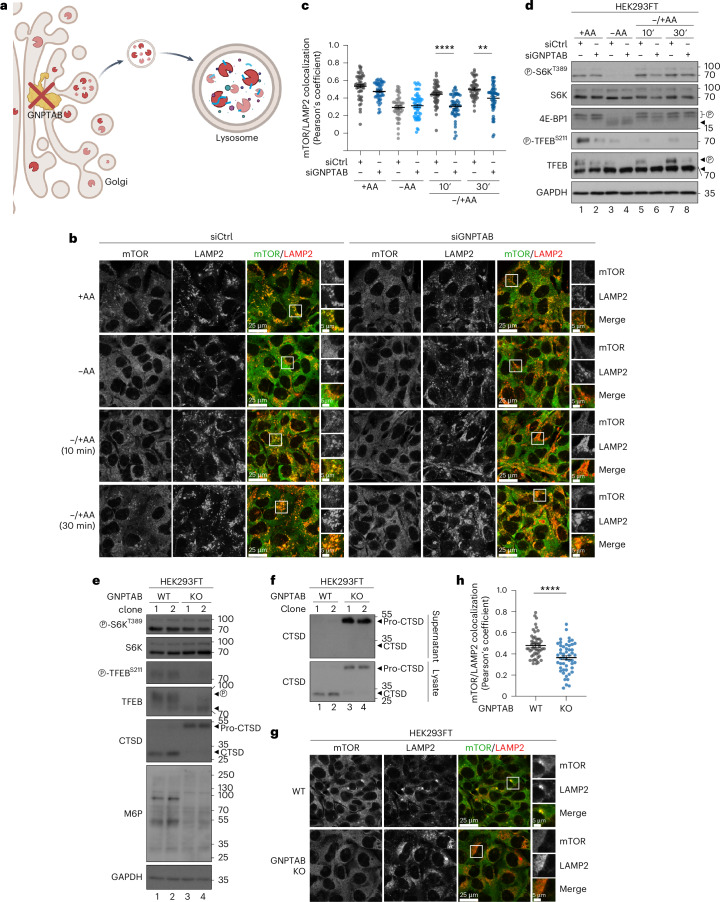
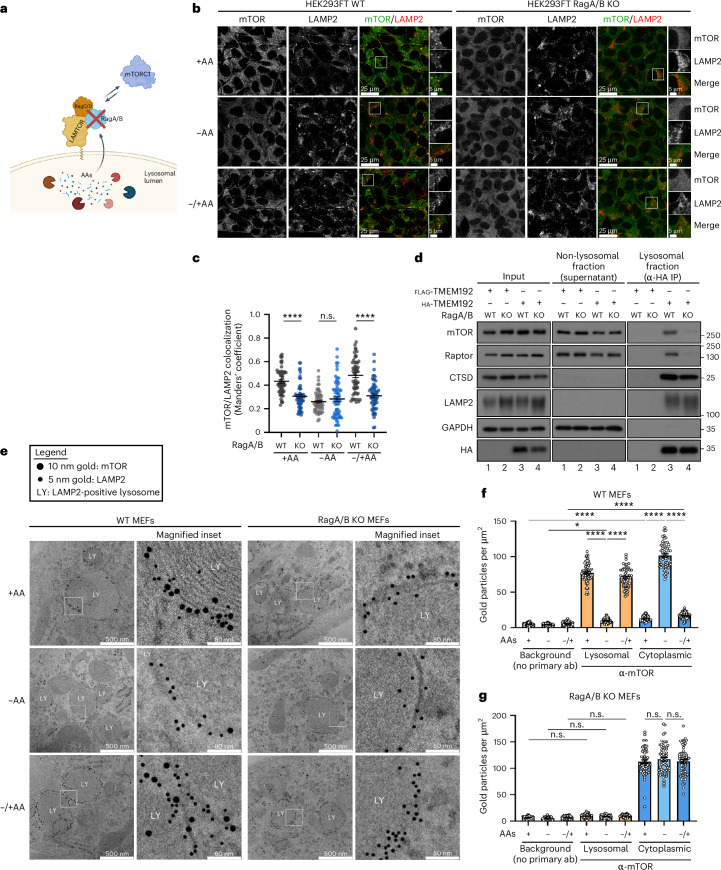
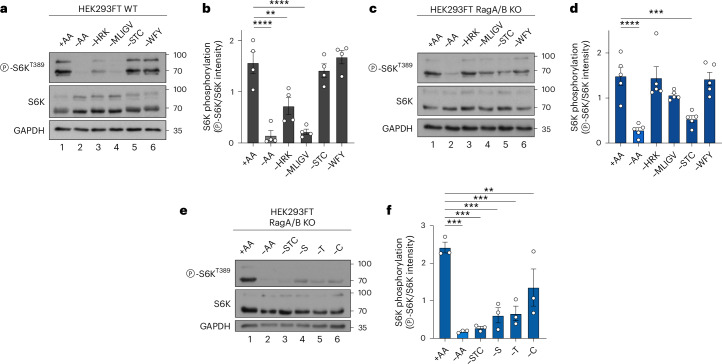
Amino acid (AA) availability is a robust determinant of cell growth through controlling mechanistic/mammalian target of rapamycin complex 1 (mTORC1) activity. According to the predominant model in the field, AA sufficiency drives the recruitment and activation of mTORC1 on the lysosomal surface by the heterodimeric Rag GTPases, from where it coordinates the majority of cellular processes. Importantly, however, the teleonomy of the proposed lysosomal regulation of mTORC1 and where mTORC1 acts on its effector proteins remain enigmatic. Here, by using multiple pharmacological and genetic means to perturb the lysosomal AA-sensing and protein recycling machineries, we describe the spatial separation of mTORC1 regulation and downstream functions in mammalian cells, with lysosomal and non-lysosomal mTORC1 phosphorylating distinct substrates in response to different AA sources. Moreover, we reveal that a fraction of mTOR localizes at lysosomes owing to basal lysosomal proteolysis that locally supplies new AAs, even in cells grown in the presence of extracellular nutrients, whereas cytoplasmic mTORC1 is regulated by exogenous AAs. Overall, our study substantially expands our knowledge about the topology of mTORC1 regulation by AAs and hints at the existence of distinct, Rag- and lysosome-independent mechanisms that control its activity at other subcellular locations. Given the importance of mTORC1 signalling and AA sensing for human ageing and disease, our findings will probably pave the way towards the identification of function-specific mTORC1 regulators and thus highlight more effective targets for drug discovery against conditions with dysregulated mTORC1 activity in the future.
氨基酸(AA)的可用性是通过控制机械/雷帕霉素靶蛋白复合物 1(mTORC1)活性来调节细胞生长的一个强有力的决定因素。根据该领域的主要模型,AA 充足会通过 Rag GTPases 异二聚体驱动 mTORC1 在溶酶体表面的募集和激活,mTORC1 从那里协调大多数细胞过程。然而,重要的是,所提出的溶酶体调节 mTORC1 的目的论和 mTORC1 在其效应蛋白上的作用位置仍然是个谜。在这里,我们通过使用多种药理学和遗传学方法来干扰溶酶体 AA 感应和蛋白质回收机制,描述了哺乳动物细胞中 mTORC1 调节和下游功能的空间分离,溶酶体和非溶酶体 mTORC1 对不同的 AA 来源磷酸化不同的效应蛋白。此外,我们揭示了由于基础溶酶体蛋白水解导致的 mTOR 的一部分定位于溶酶体中,溶酶体蛋白水解会局部提供新的 AA,即使在细胞生长在细胞外营养存在的情况下,细胞质 mTORC1 也受到外源性 AA 的调节。总的来说,我们的研究大大扩展了我们对 AA 调节 mTORC1 拓扑结构的认识,并暗示了存在其他不同的、Rag 和溶酶体独立的机制,可以控制其在其他亚细胞位置的活性。鉴于 mTORC1 信号和 AA 感应对人类衰老和疾病的重要性,我们的发现可能会为鉴定功能特异性 mTORC1 调节剂铺平道路,从而为未来针对 mTORC1 活性失调的疾病发现更有效的药物靶点提供帮助。