Licheva Mariya, Pflaum Jeremy, Babic Riccardo, Mancilla Hector, Elsässer Jana, Boyle Emily, Hollenstein David M, Jimenez-Niebla Jorge, Pleyer Jonas, Heinrich Mio, Wieland Franz-Georg, Brenneisen Joachim, Eickhorst Christopher, Brenner Johann, Jiang Shan, Hartl Markus, Welsch Sonja, Hunte Carola, Timmer Jens, Wilfling Florian, Kraft Claudine
Institute of Biochemistry and Molecular Biology, ZBMZ, Faculty of Medicine, University of Freiburg, Freiburg, Germany.
Faculty of Biology, University of Freiburg, Freiburg, Germany.
Nat Cell Biol. 2025 Feb;27(2):283-297. doi: 10.1038/s41556-024-01572-y. Epub 2025 Jan 7.
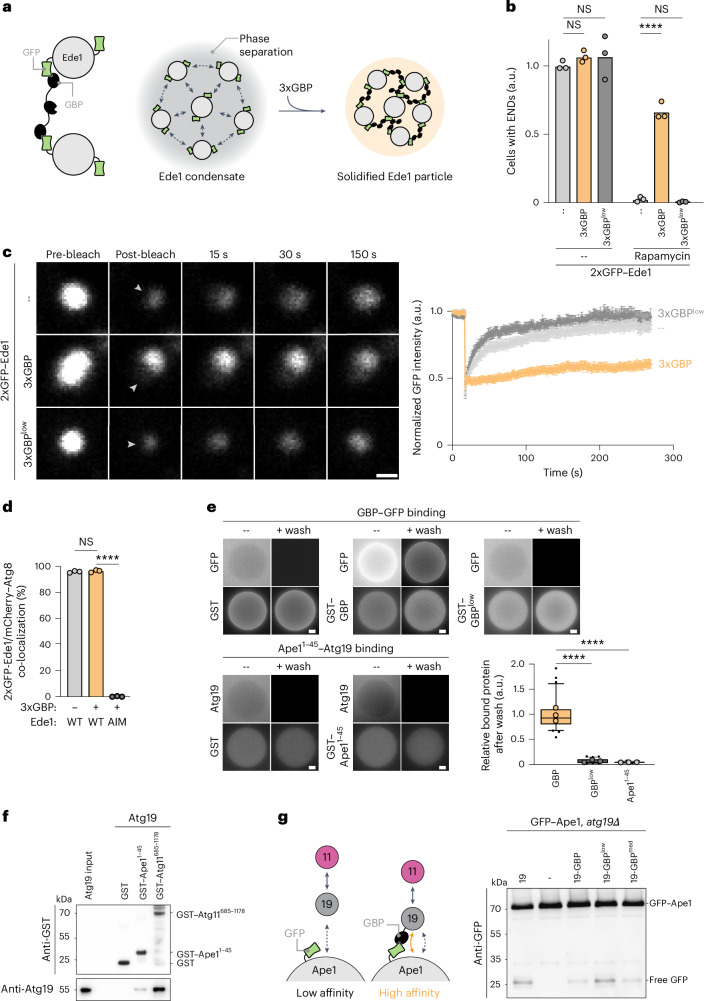
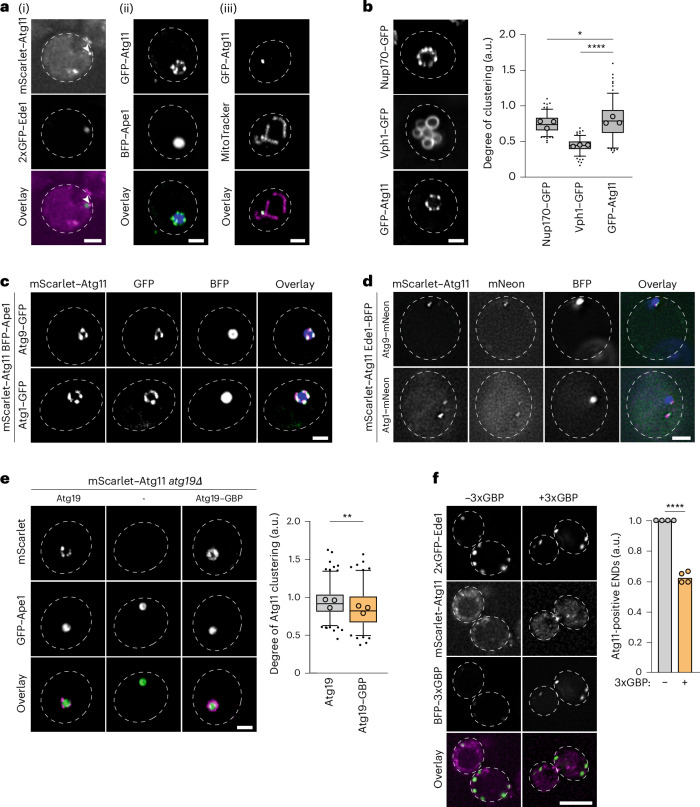
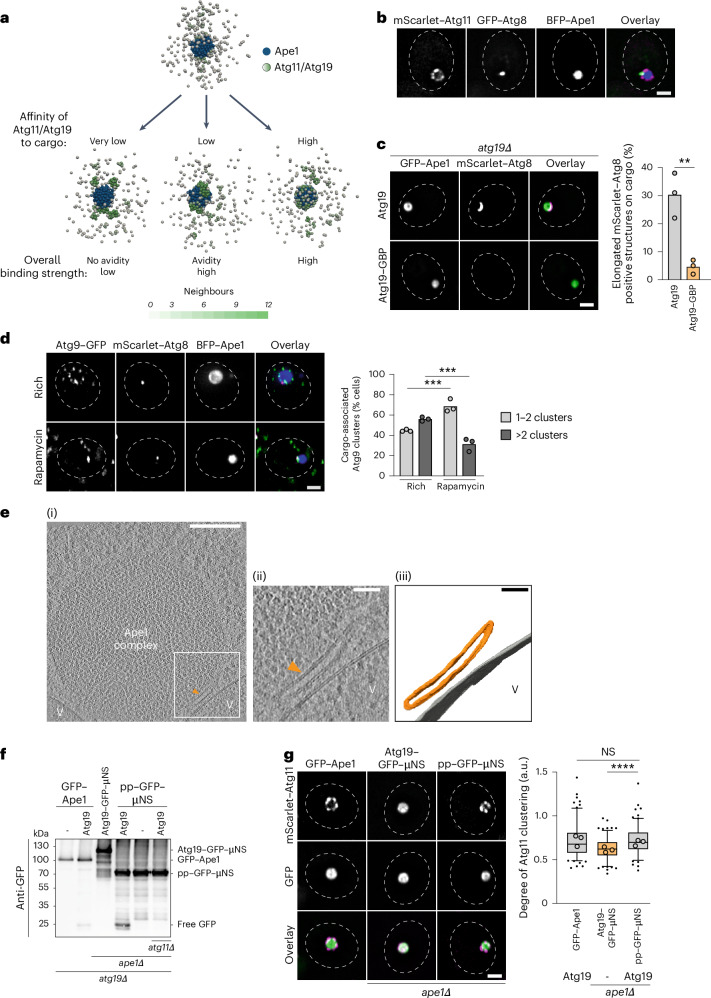
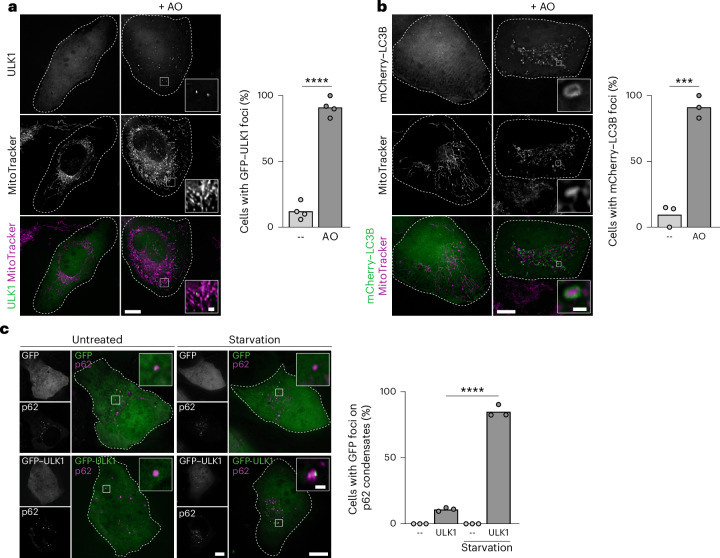
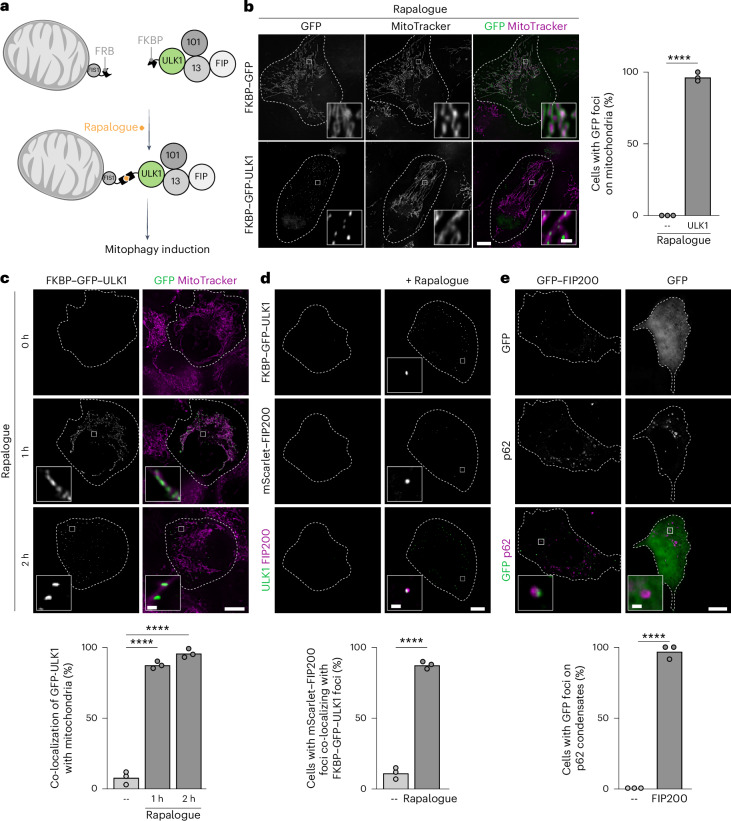
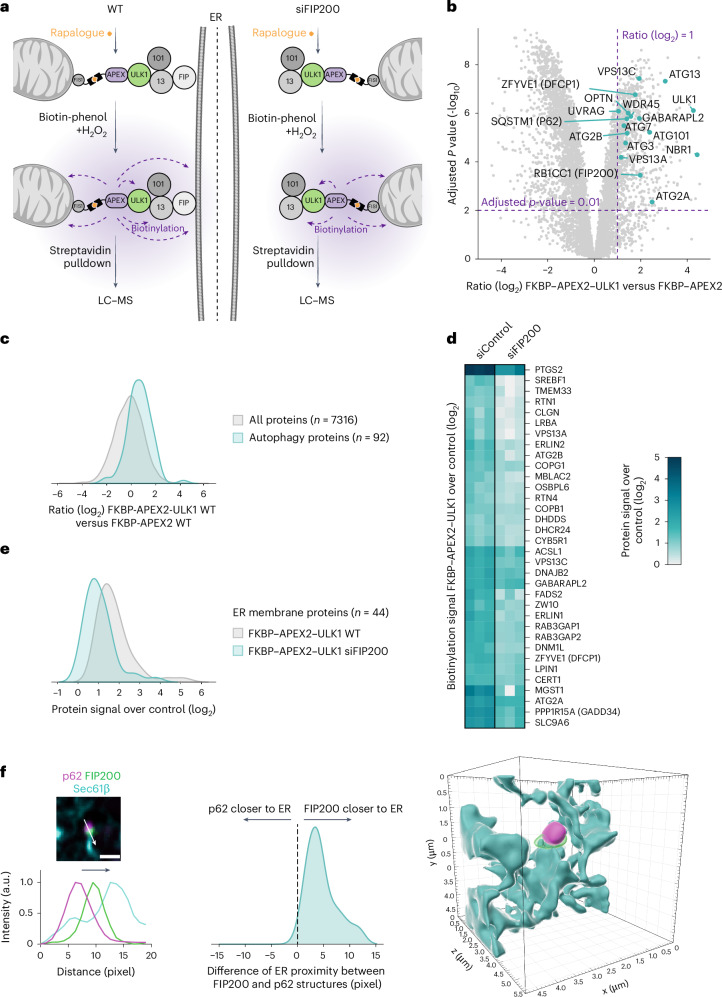
Autophagy is a key cellular quality control mechanism. Nutrient stress triggers bulk autophagy, which nonselectively degrades cytoplasmic material upon formation and liquid-liquid phase separation of the autophagy-related gene 1 (Atg1) complex. In contrast, selective autophagy eliminates protein aggregates, damaged organelles and other cargoes that are targeted by an autophagy receptor. Phase separation of cargo has been observed, but its regulation and impact on selective autophagy are poorly understood. Here, we find that key autophagy biogenesis factors phase separate into initiation hubs at cargo surfaces in yeast, subsequently maturing into sites that drive phagophore nucleation. This phase separation is dependent on multivalent, low-affinity interactions between autophagy receptors and cargo, creating a dynamic cargo surface. Notably, high-affinity interactions between autophagy receptors and cargo complexes block initiation hub formation and autophagy progression. Using these principles, we converted the mammalian reovirus nonstructural protein µNS, which accumulates as particles in the yeast cytoplasm that are not degraded, into a neo-cargo that is degraded by selective autophagy. We show that initiation hubs also form on the surface of different cargoes in human cells and are key to establish the connection to the endoplasmic reticulum, where the phagophore assembly site is formed to initiate phagophore biogenesis. Overall, our findings suggest that regulated phase separation underscores the initiation of both bulk and selective autophagy in evolutionarily diverse organisms.
自噬是一种关键的细胞质量控制机制。营养应激会触发巨自噬,在自噬相关基因1(Atg1)复合物形成并发生液-液相分离时,巨自噬会非选择性地降解细胞质物质。相比之下,选择性自噬则会清除蛋白聚集体、受损细胞器和其他被自噬受体靶向的货物。已观察到货物的相分离,但其调控以及对选择性自噬的影响仍知之甚少。在此,我们发现关键的自噬生物发生因子在酵母中会在货物表面相分离形成起始中心,随后成熟为驱动吞噬泡成核的位点。这种相分离依赖于自噬受体与货物之间多价、低亲和力的相互作用,从而形成一个动态的货物表面。值得注意的是,自噬受体与货物复合物之间的高亲和力相互作用会阻碍起始中心的形成和自噬进程。利用这些原理,我们将哺乳动物呼肠孤病毒非结构蛋白µNS(其在酵母细胞质中以颗粒形式积累且不会被降解)转化为一种可通过选择性自噬被降解的新货物。我们表明,起始中心也会在人类细胞中不同货物的表面形成,并且对于建立与内质网的连接至关重要,内质网是形成吞噬泡组装位点以启动吞噬泡生物发生的地方。总体而言,我们的研究结果表明,受调控的相分离是进化上不同生物体中巨自噬和选择性自噬起始的基础。