McLean Florence E, Omondi Brian R, Diallo Nouhoum, Otoboh Stanley, Kifude Carolyne, Abdi Abdirahman I, Lim Rivka, Otto Thomas D, Ghumra Ashfaq, Rowe J Alexandra
Institute of Immunology and Infection Research, School of Biological Sciences, University of Edinburgh, Edinburgh, United Kingdom.
KEMRI-Wellcome Trust Research Programme: Centre for Geographic Medicine Research Coast, Kilifi, Kenya.
PLoS Pathog. 2025 Jan 13;21(1):e1012434. doi: 10.1371/journal.ppat.1012434. eCollection 2025 Jan.
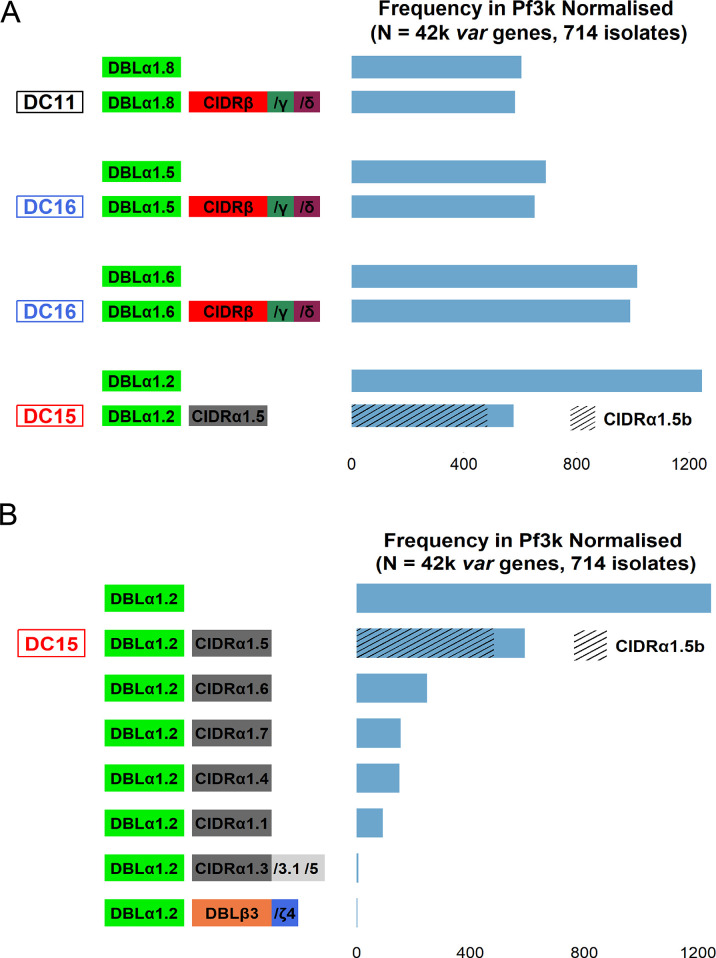
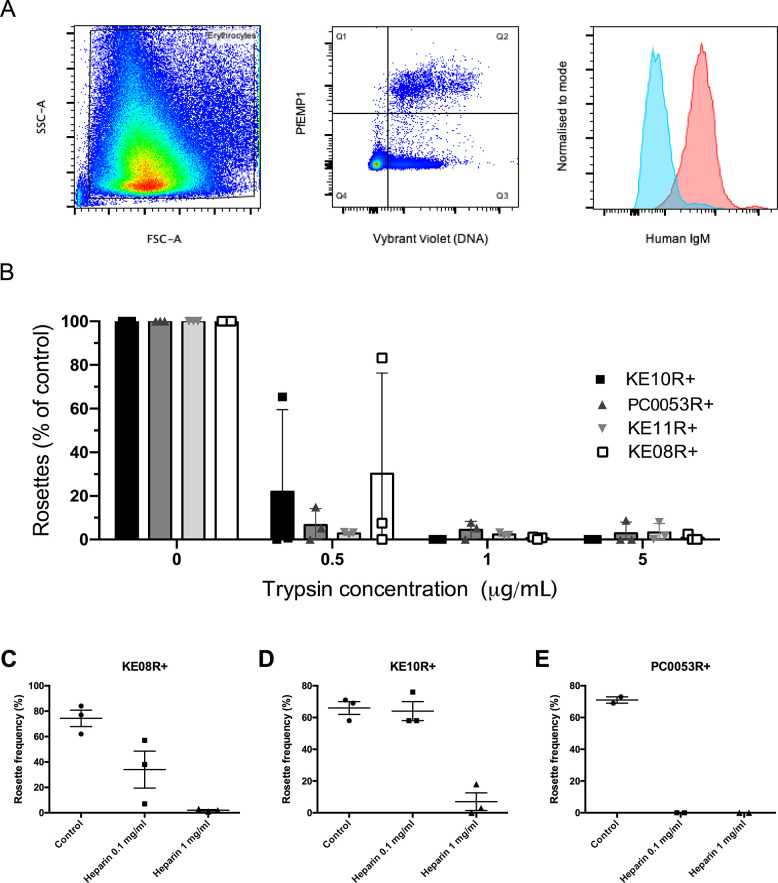
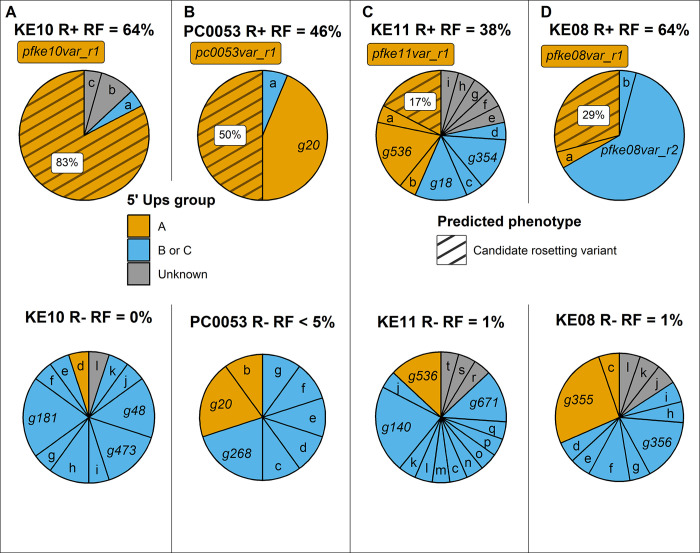
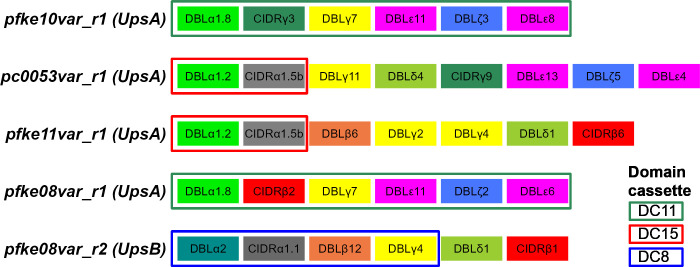
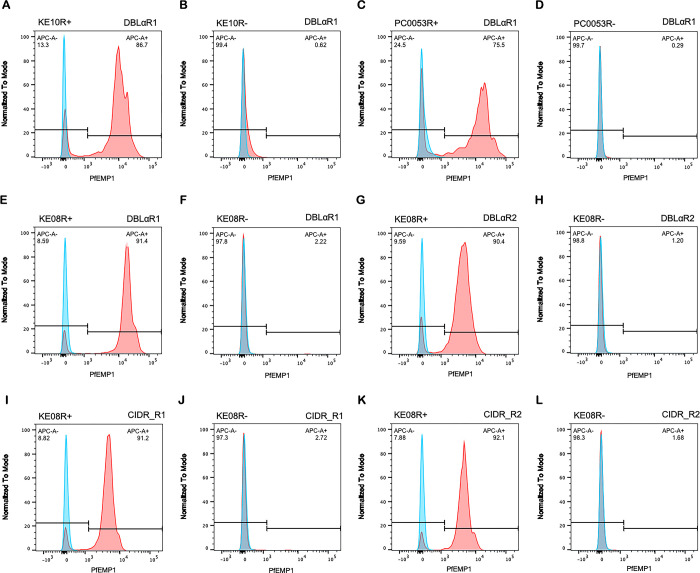
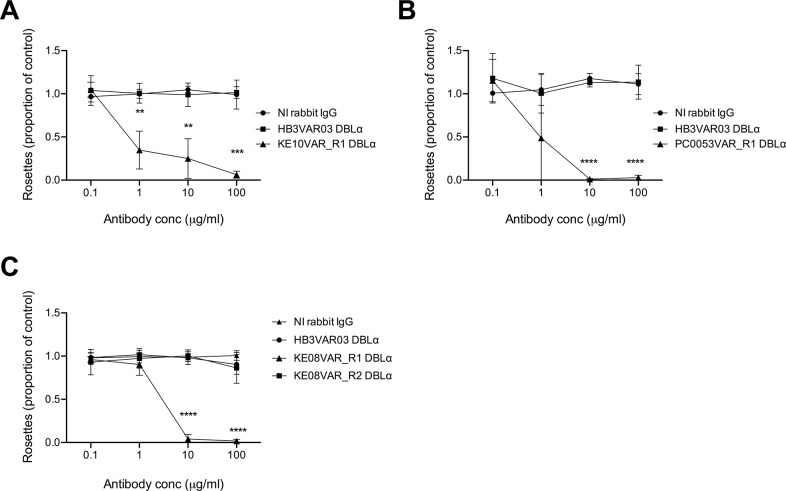
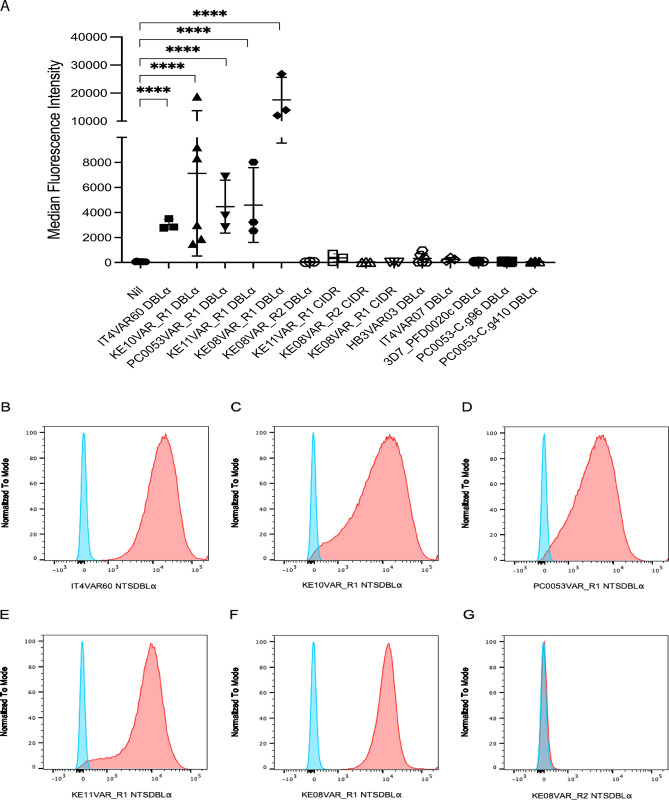
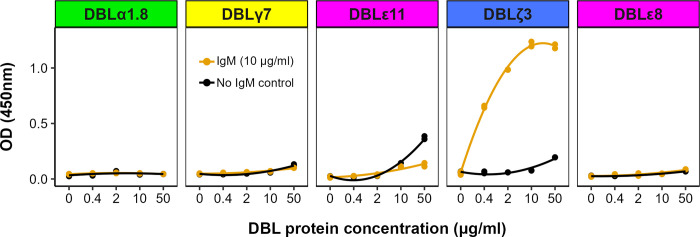
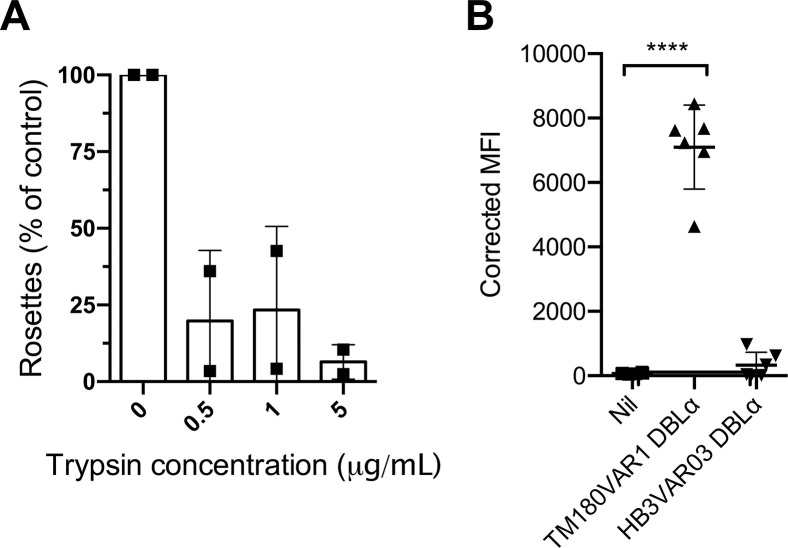
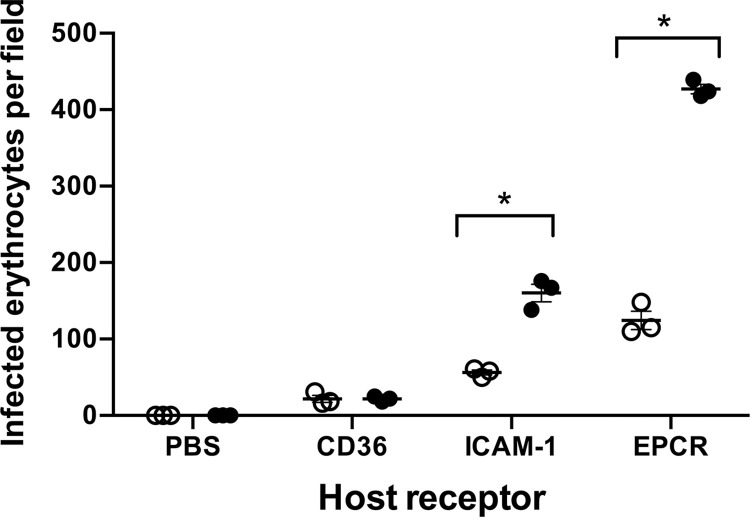
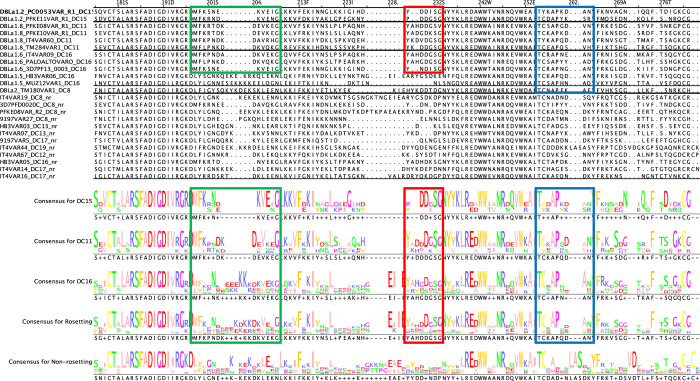
Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) is a diverse family of variant surface antigens, encoded by var genes, that mediates binding of infected erythrocytes to human cells and plays a key role in parasite immune evasion and malaria pathology. The increased availability of parasite genome sequence data has revolutionised the study of PfEMP1 diversity across multiple P. falciparum isolates. However, making functional sense of genomic data relies on the ability to infer binding phenotype from var gene sequence. For P. falciparum rosetting, the binding of infected erythrocytes to uninfected erythrocytes, the analysis of var gene/PfEMP1 sequences encoding the phenotype is limited, with only eight rosette-mediating PfEMP1 variants described to date. These known rosetting PfEMP1 variants fall into two types, characterised by N-terminal domains known as "domain cassette" 11 (DC11) and DC16. Here we test the hypothesis that DC11 and DC16 are the only PfEMP1 types in the P. falciparum genome that mediate rosetting, by examining a set of thirteen recent culture-adapted Kenyan parasite lines. We first analysed the var gene/PfEMP1 repertoires of the Kenyan lines and identified an average of three DC11 or DC16 PfEMP1 variants per genotype. In vitro rosette selection of the parasite lines yielded four with a high rosette frequency, and analysis of their var gene transcription, infected erythrocyte PfEMP1 surface expression, rosette disruption and erythrocyte binding function identified four novel rosette-mediating PfEMP1 variants. Two of these were of the predicted DC11 type (one showing the dual rosetting/IgM-Fc-binding phenotype), whereas two contained DC15 (DBLα1.2-CIDRα1.5b) a PfEMP1 type not previously associated with rosetting. We also showed that a Thai parasite line expressing a DC8-like PfEMP1 binds to erythrocytes to form rosettes. Hence, these data expand current knowledge of rosetting mechanisms and emphasize that the PfEMP1 types mediating rosetting are more diverse than previously recognised.
恶性疟原虫红细胞膜蛋白1(PfEMP1)是一个由var基因编码的多样的可变表面抗原家族,它介导受感染红细胞与人类细胞的结合,并在寄生虫免疫逃避和疟疾病理过程中起关键作用。寄生虫基因组序列数据可用性的增加彻底改变了对多个恶性疟原虫分离株中PfEMP1多样性的研究。然而,要从功能上理解基因组数据,依赖于从var基因序列推断结合表型的能力。对于恶性疟原虫的玫瑰花结形成,即受感染红细胞与未感染红细胞的结合,对编码该表型的var基因/PfEMP1序列的分析有限,迄今为止仅描述了8种介导玫瑰花结形成的PfEMP1变体。这些已知的介导玫瑰花结形成的PfEMP1变体分为两种类型,其特征是具有称为“结构域盒”11(DC11)和DC16的N端结构域。在这里,我们通过检查一组最近适应培养的肯尼亚寄生虫株系,来检验DC11和DC16是恶性疟原虫基因组中仅有的介导玫瑰花结形成的PfEMP1类型这一假设。我们首先分析了肯尼亚株系的var基因/PfEMP1库,发现每个基因型平均有三种DC11或DC16 PfEMP1变体。对这些寄生虫株系进行体外玫瑰花结选择,得到了四个玫瑰花结频率高的株系,对它们的var基因转录、受感染红细胞PfEMP1表面表达、玫瑰花结破坏和红细胞结合功能的分析,鉴定出了四种新的介导玫瑰花结形成的PfEMP1变体。其中两种属于预测的DC11类型(一种表现出双重玫瑰花结形成/IgM-Fc结合表型),而另外两种含有DC15(DBLα1.2-CIDRα1.5b),这是一种以前未与玫瑰花结形成相关联的PfEMP1类型。我们还表明,表达类似DC8的PfEMP1的泰国寄生虫株系与红细胞结合形成玫瑰花结。因此,这些数据扩展了目前对玫瑰花结形成机制的认识,并强调介导玫瑰花结形成的PfEMP1类型比以前认识的更加多样。