Lichauco Carmina, Foss Eric J, Gatbonton-Schwager Tonibelle, Athow Nelson F, Lofts Brandon, Acob Robin, Taylor Erin, Marquez James J, Lao Uyen, Miles Shawna, Bedalov Antonio
Translational Science and Therapeutics Division, Human Biology Division, Fred Hutchinson Cancer Center, Seattle, United States.
Department of Biochemistry and Department of Medicine, University of Washington, Seattle, United States.
Elife. 2025 Jan 20;13:RP97438. doi: 10.7554/eLife.97438.
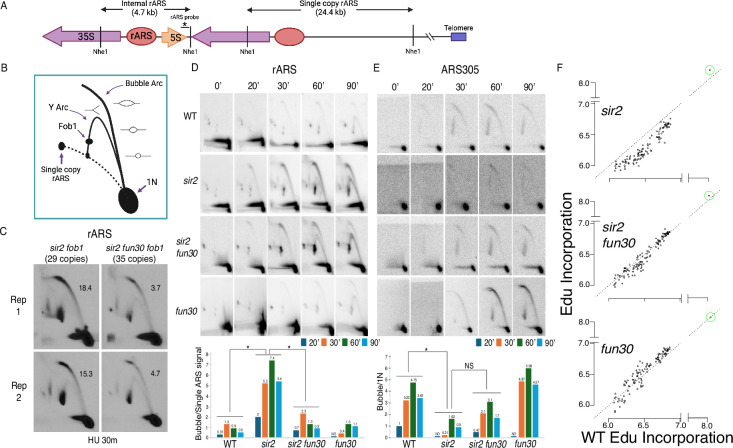
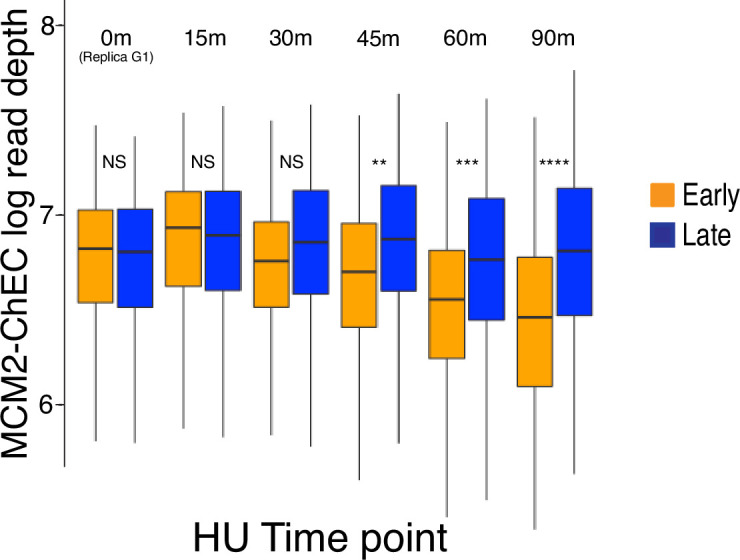
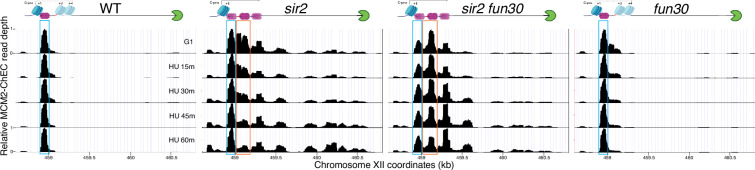
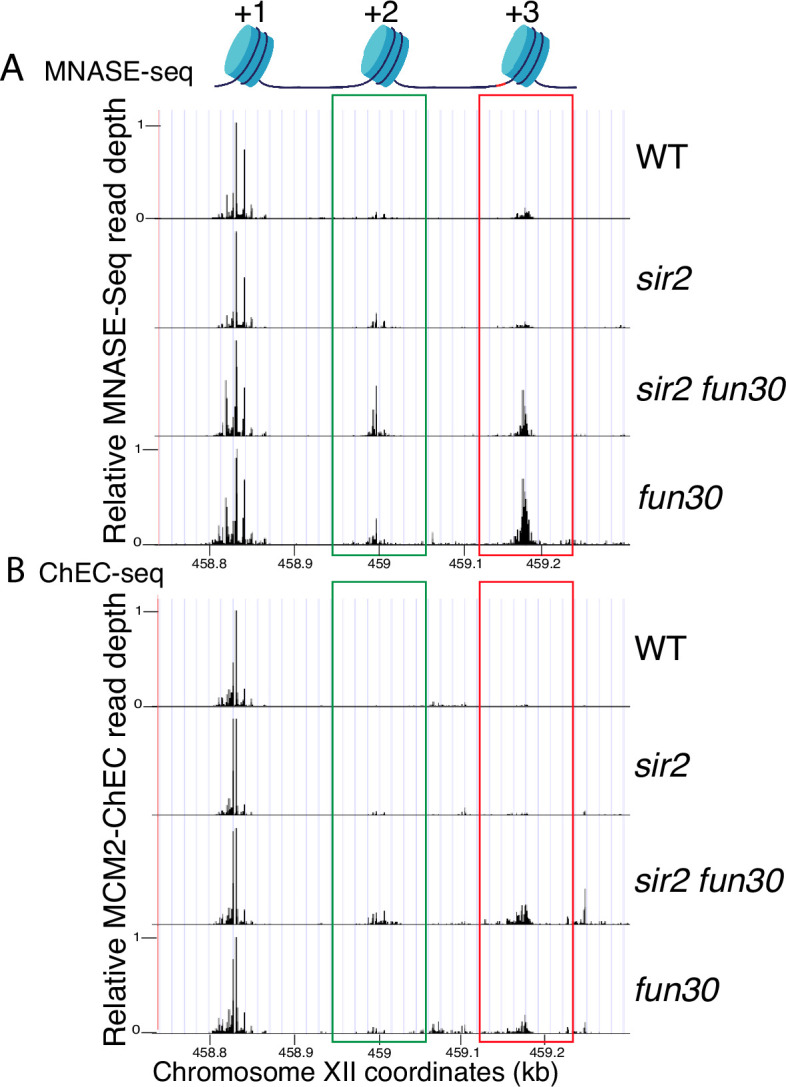
The association between late replication timing and low transcription rates in eukaryotic heterochromatin is well known, yet the specific mechanisms underlying this link remain uncertain. In , the histone deacetylase Sir2 is required for both transcriptional silencing and late replication at the repetitive ribosomal DNA (rDNA) arrays. We have previously reported that in the absence of , a de-repressed RNA PolII repositions MCM replicative helicases from their loading site at the ribosomal origin, where they abut well-positioned, high-occupancy nucleosomes, to an adjacent region with lower nucleosome occupancy. By developing a method that can distinguish activation of closely spaced MCM complexes, here we show that the displaced MCMs at rDNA origins have increased firing propensity compared to the nondisplaced MCMs. Furthermore, we found that both activation of the repositioned MCMs and low occupancy of the adjacent nucleosomes critically depend on the chromatin remodeling activity of . Our study elucidates the mechanism by which Sir2 delays replication timing, and it demonstrates, for the first time, that activation of a specific replication origin in vivo relies on the nucleosome context shaped by a single chromatin remodeler.
真核生物异染色质中复制时间较晚与转录速率较低之间的关联是众所周知的,但这种联系背后的具体机制仍不确定。在酿酒酵母中,组蛋白脱乙酰酶Sir2对于重复核糖体DNA(rDNA)阵列处的转录沉默和复制延迟都是必需的。我们之前报道过,在缺乏Sir2的情况下,去抑制的RNA聚合酶II将MCM复制解旋酶从其在核糖体起源处的加载位点重新定位,在该位点它们紧邻定位良好、占有率高的核小体,转移至相邻的核小体占有率较低的区域。通过开发一种能够区分紧密间隔的MCM复合物激活的方法,我们在此表明,与未移位的MCM相比,rDNA起源处移位的MCM具有更高的起始倾向。此外,我们发现重新定位的MCM的激活以及相邻核小体的低占有率都严重依赖于Sir2的染色质重塑活性。我们的研究阐明了Sir2延迟复制时间的机制,并且首次证明了体内特定复制起点的激活依赖于由单个染色质重塑因子塑造的核小体环境。