Lin Yan, Liu Bingjie, Hu Yuxiang, Li Ganghua, Liu Zhenghui, Ding Yanfeng, Chen Lin
College of Agriculture, Nanjing Agricultural University, Nanjing, 210095, China.
Key Laboratory of Crop Physiology and Ecology in Southern China, Ministry of Agricultural University, Nanjing, 210095, China.
Rice (N Y). 2025 Jun 19;18(1):54. doi: 10.1186/s12284-025-00816-1.
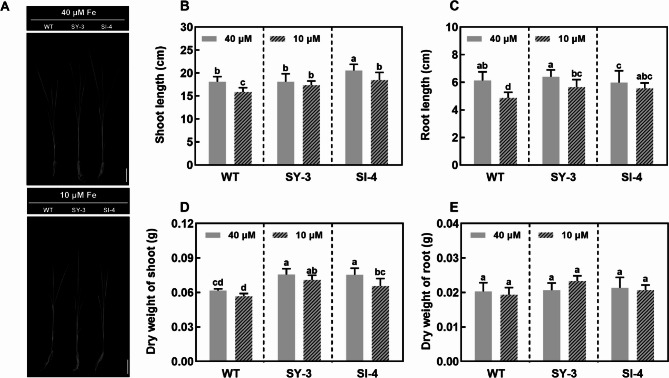
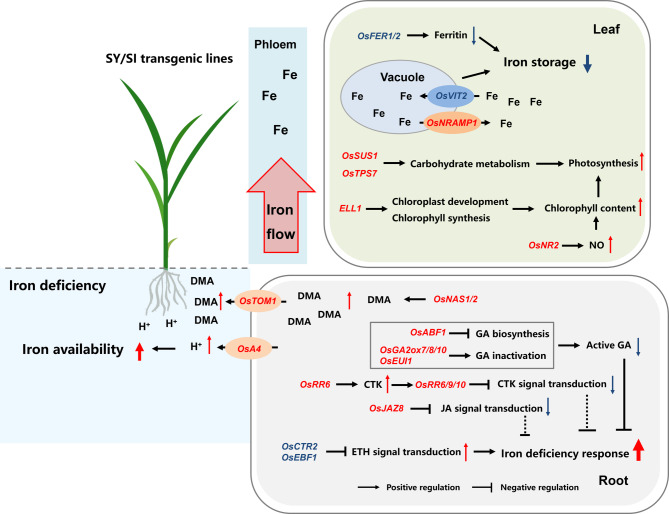
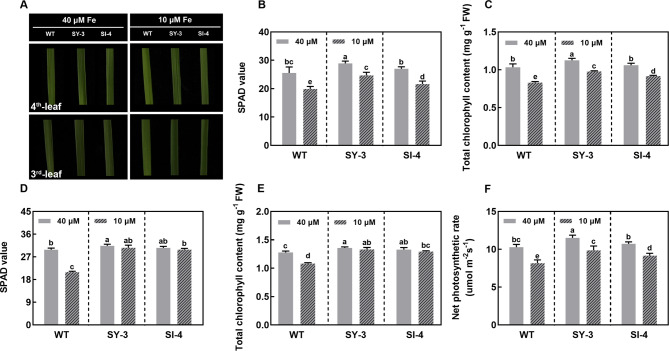
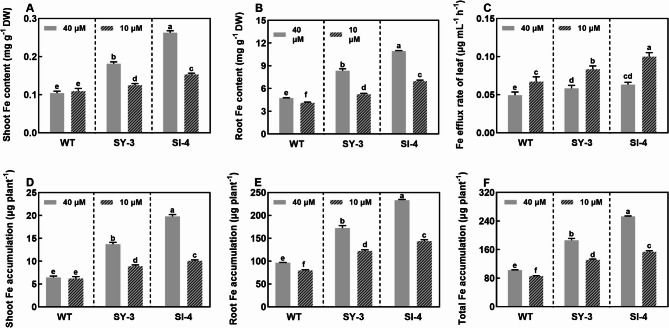
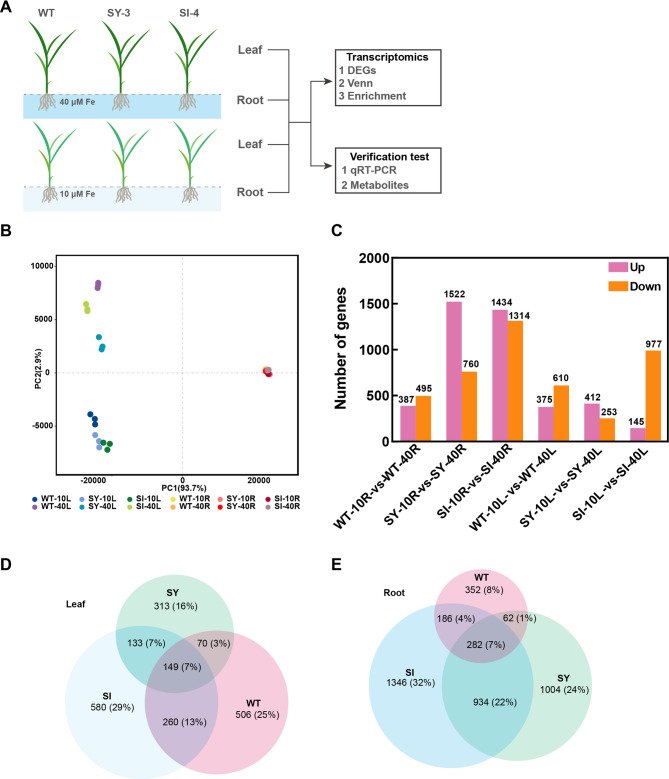
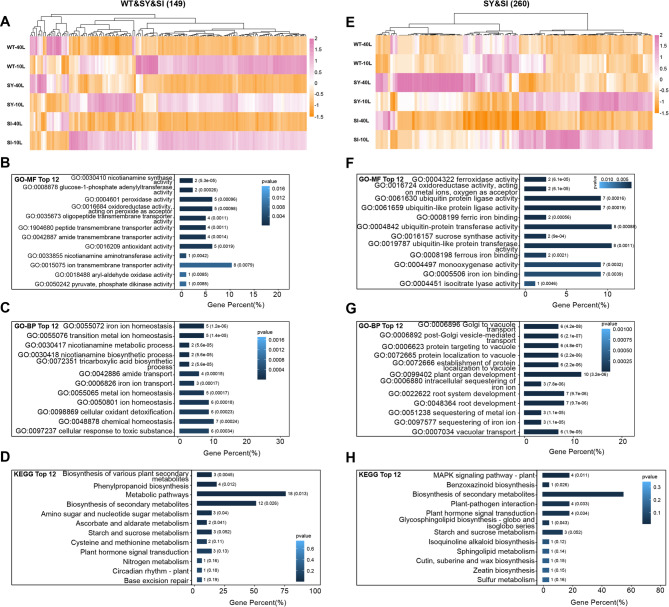
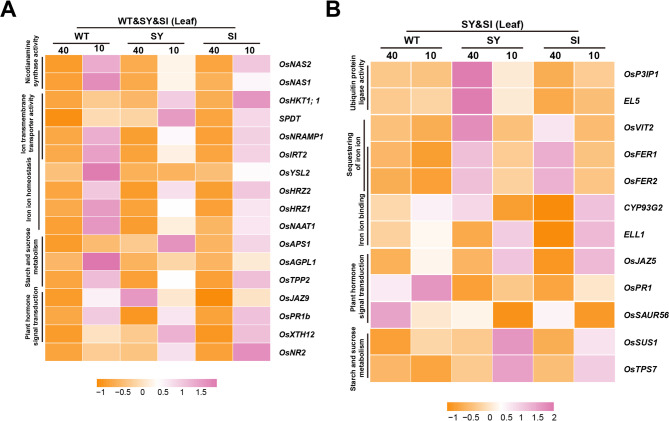
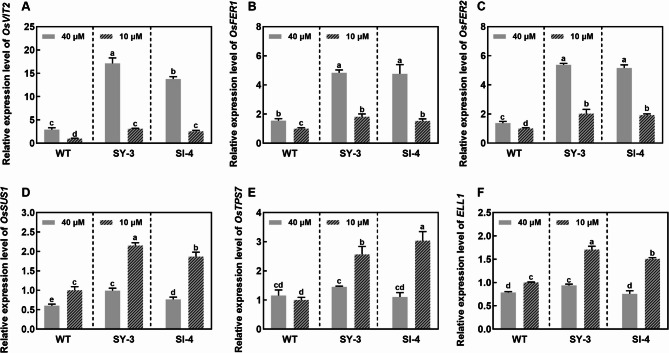
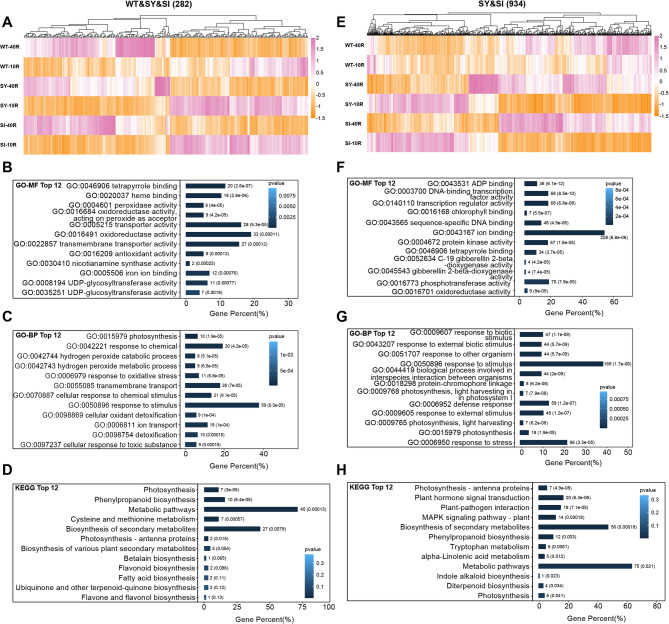
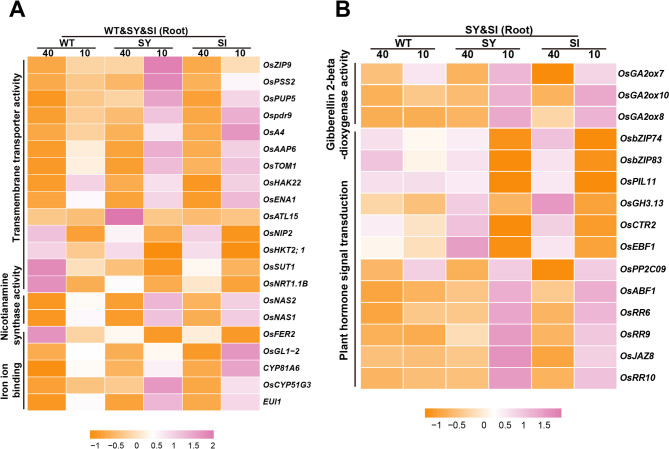
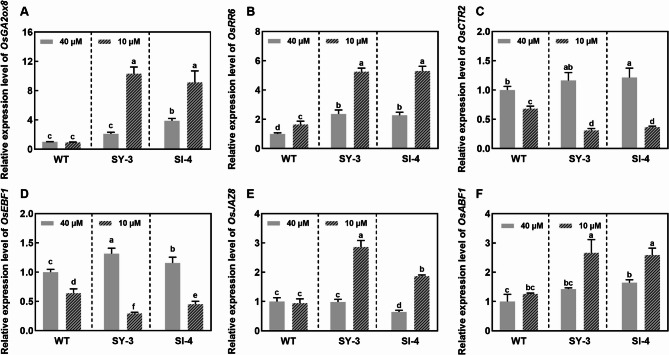
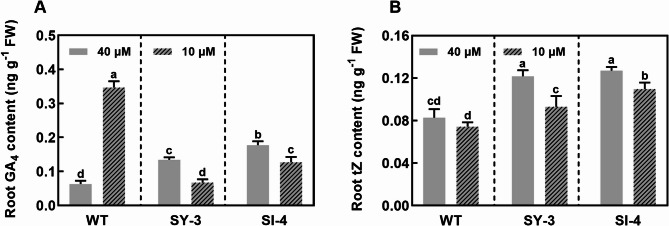
Iron (Fe) is essential for normal plant growth and development. In rice, Fe deficiency leads to stunted growth, leaf chlorosis, reduced photosynthetic capacity, and ultimately, yield loss. Most studies have focused on investigating the mechanisms of Fe deficiency responses in rice roots; however, the effects of shoot Fe redistribution on Fe deficiency response remain poorly understood. Phloem transport plays a vital role in distributing Fe to new tissues. To investigate the effects of enhanced phloem-mediated Fe transport on rice adaptability to iron deficiency, we subjected transgenic lines with higher phloem Fe efflux rates and wild-type (WT) plants to Fe-deficient conditions. The growth, leaf photosynthetic rate, and Fe content of transgenic and WT seedlings under different Fe concentrations were compared. The results showed that the transgenic lines exhibited elevated shoot length, root length, shoot dry weight, leaf chlorophyll content, and net photosynthetic rates under Fe-deficient conditions. Under both Fe-sufficient and Fe-deficient conditions, the transgenic lines had significantly higher Fe content, Fe accumulation, and phloem Fe efflux rates than the WT. RNA sequencing (RNA-seq) analysis revealed that enhanced Fe transport via phloem resulted in improved Fe availability through the sequestration of Fe ions and vacuolar transport pathways in the shoots. It also upregulated the EARLY LESION LEAF 1 (ELL1) expression and modulated the sucrose synthase activity, thereby promoting chlorophyll synthesis and leaf photosynthesis. Additionally, enhanced Fe transport influenced the gibberellin (GA) catabolism and plant hormone signal transduction in the roots, reducing the GA content and modulating the cytokinin (CTK), jasmonic acid (JA), and ethylene (ETH) signaling to induce Fe deficiency response and promote Fe uptake. These findings demonstrate that phloem-mediated Fe transport participated in Fe deficiency response, and enhancing this improved the adaptability of rice seedlings to low Fe conditions. In specific, rice seedlings with a high capacity for phloem-mediated Fe transport exhibited a strong iron uptake, translocation, and remobilization capacity, thereby maintaining normal growth and development and successfully adapting to the low-Fe environment.
铁(Fe)对于植物的正常生长和发育至关重要。在水稻中,缺铁会导致生长发育迟缓、叶片黄化、光合能力下降,最终造成产量损失。大多数研究都集中在探究水稻根系缺铁响应的机制;然而,地上部铁再分配对缺铁响应的影响仍知之甚少。韧皮部运输在将铁分配到新组织中起着至关重要的作用。为了研究增强韧皮部介导的铁运输对水稻缺铁适应性的影响,我们将具有较高韧皮部铁外排率的转基因株系和野生型(WT)植株置于缺铁条件下。比较了不同铁浓度下转基因和WT幼苗的生长、叶片光合速率和铁含量。结果表明,在缺铁条件下,转基因株系的地上部长度、根长、地上部干重、叶片叶绿素含量和净光合速率均有所提高。在铁充足和缺铁条件下,转基因株系的铁含量、铁积累量和韧皮部铁外排率均显著高于WT。RNA测序(RNA-seq)分析表明,通过韧皮部增强铁运输可通过螯合地上部的铁离子和液泡运输途径来提高铁的有效性。它还上调了早期损伤叶片1(ELL1)的表达并调节蔗糖合酶活性,从而促进叶绿素合成和叶片光合作用。此外,增强的铁运输影响了根中的赤霉素(GA)分解代谢和植物激素信号转导,降低了GA含量并调节了细胞分裂素(CTK)、茉莉酸(JA)和乙烯(ETH)信号,以诱导缺铁响应并促进铁吸收。这些发现表明,韧皮部介导的铁运输参与了缺铁响应,增强这一过程可提高水稻幼苗对低铁条件的适应性。具体而言,具有高韧皮部介导铁运输能力的水稻幼苗表现出强大的铁吸收、转运和再利用能力,从而维持正常的生长发育并成功适应低铁环境。