Kress B C, Hirani S, Freeze H H, Little L, Miller A L
Biochem J. 1982 Dec 1;207(3):421-8. doi: 10.1042/bj2070421.
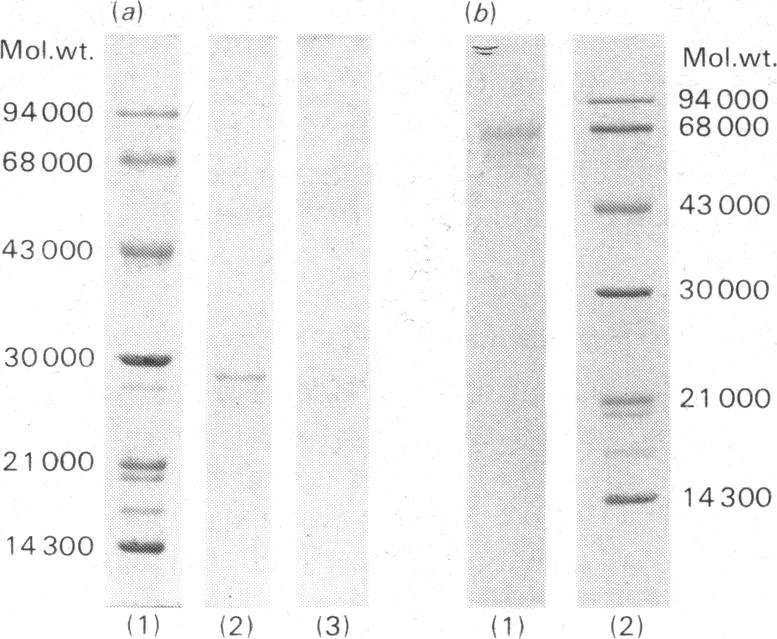
Mucolipidosis III acid hydrolases possess an altered carbohydrate recognition marker needed for their lysosomal localization. As a result of this alteration, a portion of these enzymes is secreted from the cell to the extracellular spaces. The structural changes that may have occurred to one of these secreted enzymes, beta-N-acetyl-d-hexosaminidase A (EC 3.2.1.52) were investigated. Normal and mucolipidosis III urinary beta-N-acetyl-d-hexosaminidase A were purified to apparent homogeneity by using affinity [Sepharose-2-acetamido-N-(epsilon-aminocaproyl)-2-deoxy-beta- d-glucopyranosylamine] and ion-exchange (DEAE- and CM-cellulose) chromatography. Sodium dodecyl sulphate/polyacrylamide-slab-gel electrophoresis showed that both enzymes had similar subunit patterns consisting of apparent mol.wts. of 68000, 60000-58000, 55000 and 29000. Differences, however, were noted in the relative proportions of the protein bands where the normal urinary beta-N-acetyl-d-hexosaminidase A contained predominantly the smaller subunits, whereas the mucolipidosis III enzyme had a predominance of the larger subunits. The binding of mucolipidosis III beta-N-acetyl-d-hexosaminidase A to Ricinus communis lectin and concanavalin A with and without endo-beta-N-acetyl-d-glucosaminidase H treatment indicated that the mutation leads to a modification of a portion of the normally occurring high-mannose-type oligosaccharide units to the complex-type. This was further supported by carbohydrate compositional analysis, which revealed a mannose/galactose ratio of 2.1 for the mucolipidosis III beta-N-acetyl-d-hexosaminidase A compared with a ratio of 3.5 for the normal enzyme. Our results indicate that as a result of their inability to be properly localized to the lysosome the majority of the mucolipidosis III lysosomal hydrolase high-mannose oligosaccharide units are further processed to the complex-type before secretion of predominantly higher-molecular-weight subunits from the cell.
黏脂贮积症III型酸性水解酶具有改变的碳水化合物识别标记,这是其溶酶体定位所必需的。由于这种改变,这些酶中的一部分从细胞分泌到细胞外空间。研究了这些分泌酶之一β-N-乙酰-D-己糖胺酶A(EC 3.2.1.52)可能发生的结构变化。通过使用亲和色谱法(琼脂糖-2-乙酰氨基-N-(ε-氨基己酰基)-2-脱氧-β-D-吡喃葡萄糖胺)和离子交换色谱法(DEAE-纤维素和CM-纤维素)将正常和黏脂贮积症III型尿β-N-乙酰-D-己糖胺酶A纯化至表观均一性。十二烷基硫酸钠/聚丙烯酰胺平板凝胶电泳显示,两种酶具有相似的亚基模式,由表观分子量为68000、60000 - 58000、55000和29000组成。然而,在蛋白条带的相对比例上存在差异,正常尿β-N-乙酰-D-己糖胺酶A主要包含较小的亚基,而黏脂贮积症III型酶则以较大的亚基为主。黏脂贮积症III型β-N-乙酰-D-己糖胺酶A在有无内切β-N-乙酰-D-葡糖胺酶H处理的情况下与蓖麻凝集素和伴刀豆球蛋白A的结合表明,该突变导致一部分正常存在的高甘露糖型寡糖单元转变为复合型。碳水化合物组成分析进一步支持了这一点,该分析显示黏脂贮积症III型β-N-乙酰-D-己糖胺酶A的甘露糖/半乳糖比率为2.1,而正常酶的比率为3.5。我们的结果表明,由于黏脂贮积症III型溶酶体水解酶无法正确定位于溶酶体,其大多数高甘露糖寡糖单元在从细胞分泌主要是较高分子量亚基之前进一步加工为复合型。