Natarajan V, Madden M J, Salzman N P
Proc Natl Acad Sci U S A. 1984 Oct;81(20):6290-4. doi: 10.1073/pnas.81.20.6290.
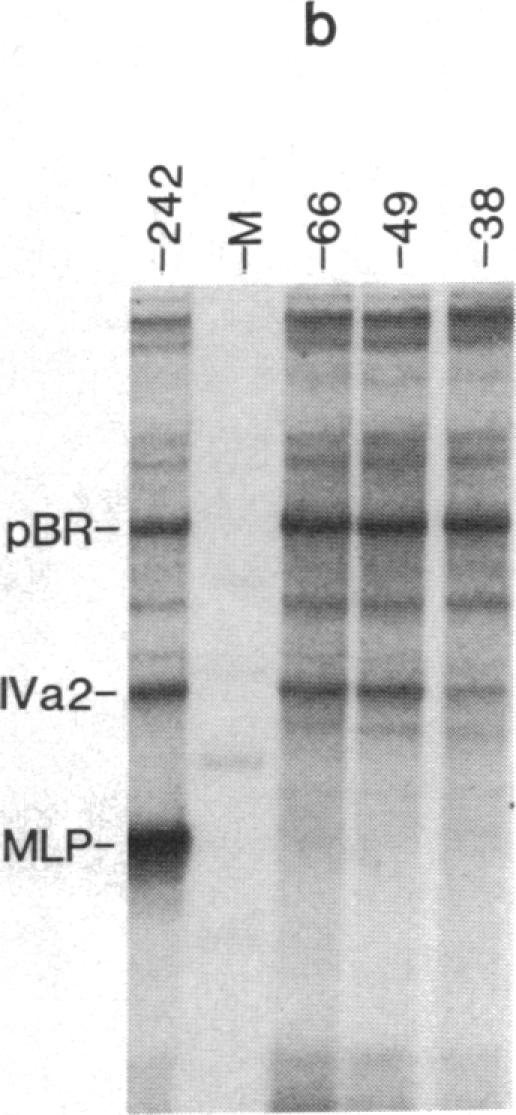
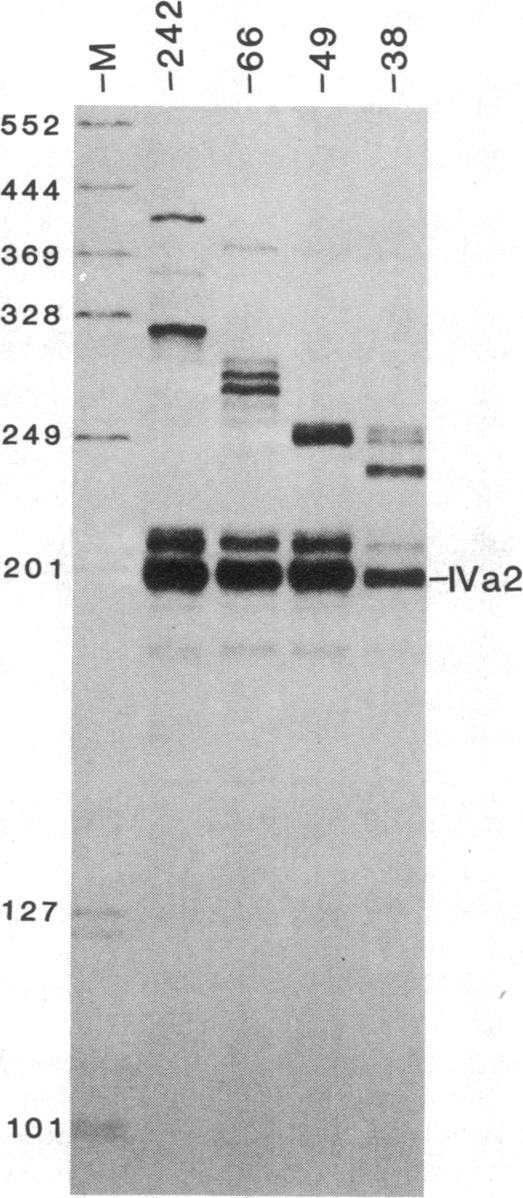
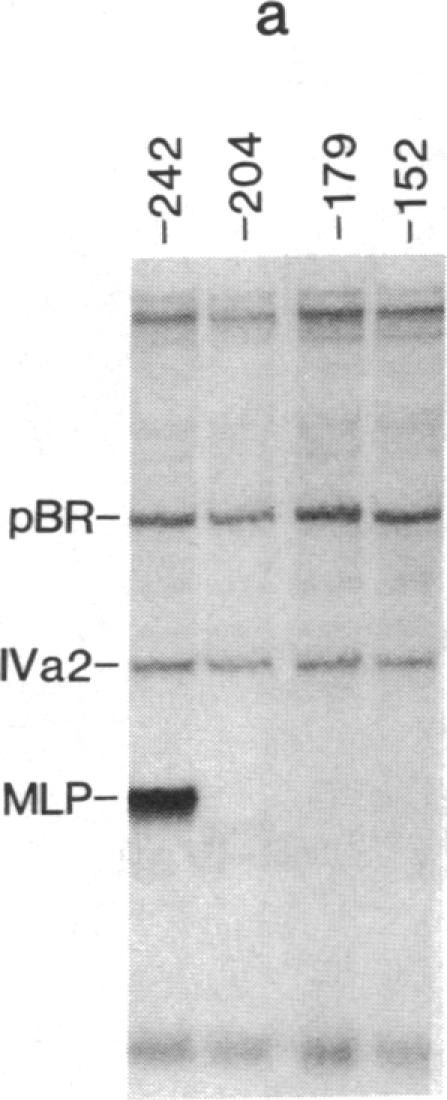
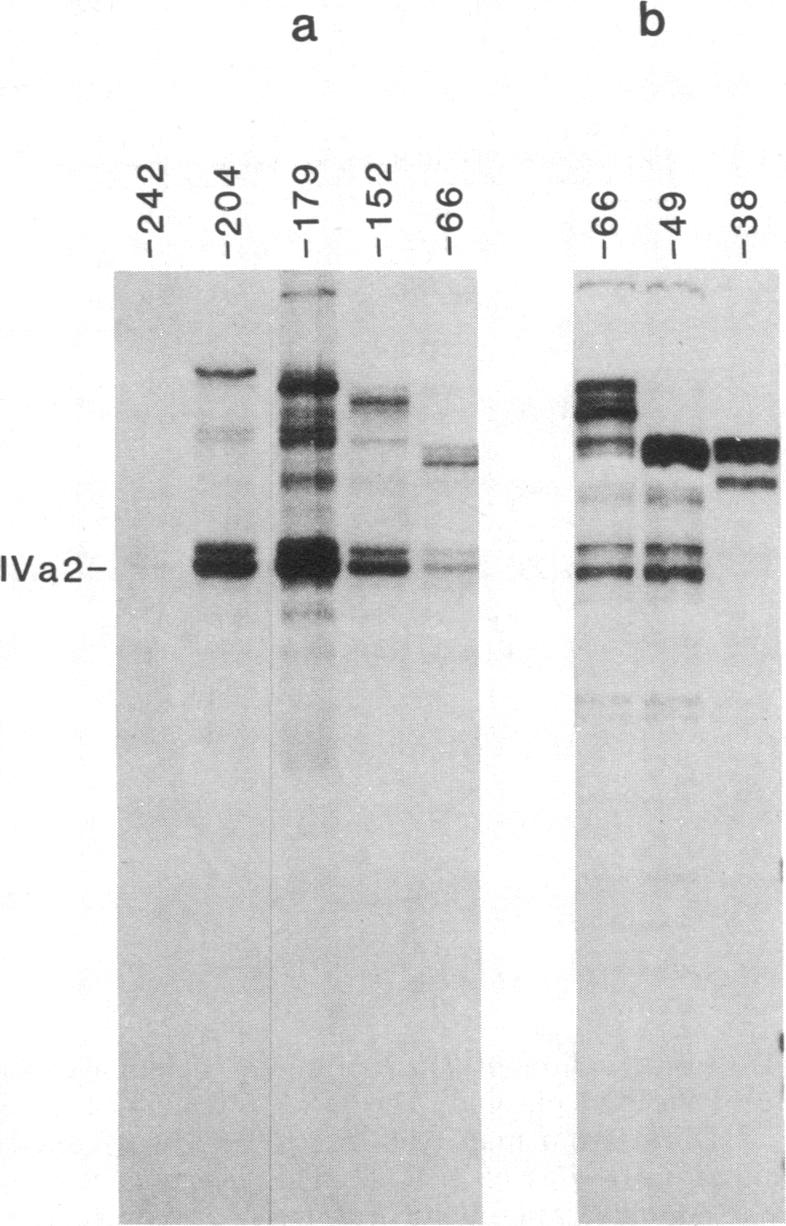
The adenovirus IVa2 gene, which is expressed at an intermediate time in the viral infectious cycle, is separated from the adenovirus major late promoter (MLP) 5' start site by 210 base pairs and is transcribed from the opposite strand. In contrast to the MLP, the IVa2 gene does not contain a "TATA" box upstream from its 5' start sites. By using a series of deletion mutants, two upstream control regions that are rich in cytidine residues, one proximal to the cap site at nucleotide positions -39 to -48 and a distal domain between nucleotide positions -152 and -242 have been identified as essential for IVa2 transcription (IVa2 cap site is nucleotide position + 1). Transcription efficiency is decreased by 70-90% after the deletion of a proximal C-rich domain when either linear or supercoiled DNAs were used as template. However, distal sequences functioned as transcriptional control domains only with covalently closed DNA templates. The deletion of both the proximal and distal regions from covalently closed DNA templates reduces the levels of IVa2 transcription by a factor of 100-150. When the plasmid pAd242 that contains the 5' start sites of adenovirus MLP and IVa2 is transcribed, there is essentially a complete suppression of transcription of the adenovirus IVa2 gene. The transcription efficiency of IVa2 is increased 10-fold after deletion of the MLP cap site. A model based on a shared entry site for RNA polymerase II and competition between major late and IVa2 promoters is proposed to explain the in vitro transcriptional results.
腺病毒IVa2基因在病毒感染周期的中期表达,与腺病毒主要晚期启动子(MLP)的5'起始位点相隔210个碱基对,并且从相反链转录。与MLP不同,IVa2基因在其5'起始位点上游不包含“TATA”框。通过使用一系列缺失突变体,已鉴定出两个富含胞嘧啶残基的上游控制区域,一个在核苷酸位置-39至-48处靠近帽位点,另一个在核苷酸位置-152和-242之间的远端结构域,它们对于IVa2转录至关重要(IVa2帽位点为核苷酸位置+1)。当使用线性或超螺旋DNA作为模板时,缺失近端富含C的结构域后转录效率降低70-90%。然而,远端序列仅与共价闭合的DNA模板一起作为转录控制结构域起作用。从共价闭合的DNA模板中删除近端和远端区域会使IVa2转录水平降低100-150倍。当转录包含腺病毒MLP和IVa2的5'起始位点的质粒pAd242时,腺病毒IVa2基因的转录基本上被完全抑制。删除MLP帽位点后,IVa2的转录效率提高了10倍。提出了一个基于RNA聚合酶II共享进入位点以及主要晚期启动子和IVa2启动子之间竞争的模型来解释体外转录结果。