Chan-Palay V, Palay S L, Wu J Y
Proc Natl Acad Sci U S A. 1982 Jul;79(13):4221-5. doi: 10.1073/pnas.79.13.4221.
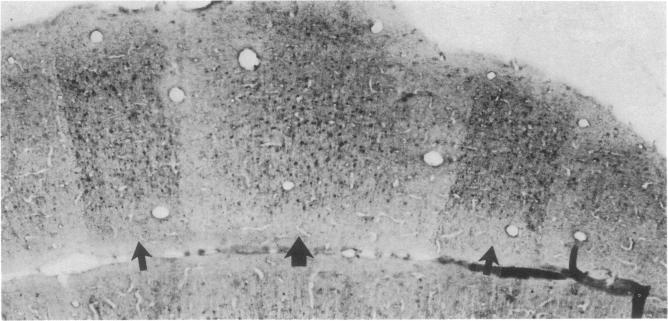
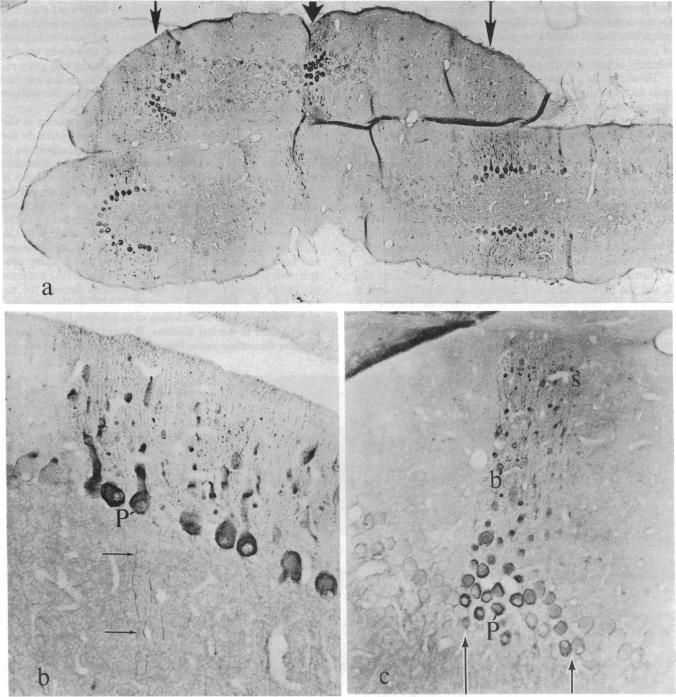
Taurine neurons in the cerebellum of rabbit, rat, and mouse were localized at the light microscope level by using polyclonal antibodies against cysteine sulfinic acid decarboxylase (CSADCase; EC 4.1.1.29), the enzyme responsible for the conversion of cysteine sulfinic acid to hypotaurine and of cysteic acid to taurine. The indirect peroxidase-antiperoxidase method was used on Vibratome sections and on serial sections of paraffin-embedded tissue. Intensification of CSADCase immunoreactivity was achieved by pretreatment of the animal with L-cysteine or L-cysteic acid intravenously 1-2 hr prior to perfusion. A combination of L-cysteic acid and demecolcine, which retards axoplasmic flow, was most effective in maximizing CSADCase immunoreactivity. Although these treatments intensified immunoreactivity in neurons, no more cells were reactive than in untreated controls. L-Glutamic acid did not increase CSADCase immunoreactivity but did increase immunoreactivity with antibodies against L-glutamic acid decarboxylase (GAD; EC 4.1.1.15), the synthetic enzyme for gamma-aminobutyric acid. Specificity was established by negative results obtained with various control incubations including the use of CSADCase antiserum preabsorbed with the antigen. Taurine neurons of the cerebellar cortex are arranged in sagittal microbands, defined by intensely CSADCase-reactive Purkinje neurons and their axons and dendrites, together with stellate, basket, and Golgi cells and their processes. In the vermis there is a narrow midline band, flanked laterally by three wider bands on either side, each separated from the next by an unreactive zone. Although the zonal borders are sharp, the interzonal areas contain some CSADCase-immunoreactive axons but no cell bodies. The seven vermal bands are best observed in the anterior lobe. Others exist in the lateral hemispheres. The paraflocculus and flocculus contain numerous intensely immunoreactive neurons, and banding is difficult to discern. Lobule X of the vermis is also heavily endowed with taurine neurons. Numerous large and medium-sized deep cerebellar and vestibular nuclei are also immunoreactive. These observations indicate that cerebellar neurons are chemically heterogeneous but that neurons of similar chemical signature in the cerebellar cortex are organized into sagittal microbands. This corroborates our earlier evidence that Purkinje cells containing motilin and those containing both motilin and gamma-aminobutyric acid are also arranged in vermal sagittal microbands. The midline vermal band contains Purkinje neurons with multiple neuroactive substances-taurine, gamma-aminobutyric acid, and motilin. It remains to be determined how this chemical zonation in the cerebellar cortex relates to the banded afferent innervation from spinal, vestibular, reticular, and olivary sources.
通过使用针对半胱氨酸亚磺酸脱羧酶(CSADCase;EC 4.1.1.29)的多克隆抗体,在光镜水平上对兔、大鼠和小鼠小脑内的牛磺酸能神经元进行定位,该酶负责将半胱氨酸亚磺酸转化为次牛磺酸以及将半胱氨酸转化为牛磺酸。间接过氧化物酶 - 抗过氧化物酶法用于振动切片机切片以及石蜡包埋组织的连续切片。在灌注前1 - 2小时通过静脉注射L - 半胱氨酸或L - 半胱氨酸对动物进行预处理,可增强CSADCase免疫反应性。L - 半胱氨酸和秋水仙碱的组合(秋水仙碱可延缓轴浆运输)在使CSADCase免疫反应性最大化方面最为有效。尽管这些处理增强了神经元中的免疫反应性,但与未处理的对照组相比,有反应的细胞数量并未增加。L - 谷氨酸并未增加CSADCase免疫反应性,但确实增加了针对L - 谷氨酸脱羧酶(GAD;EC 4.1.1.15,γ - 氨基丁酸的合成酶)抗体的免疫反应性。通过各种对照孵育获得的阴性结果确定了特异性,包括使用用抗原预吸收的CSADCase抗血清。小脑皮质的牛磺酸能神经元排列成矢状微带,由强烈CSADCase反应性的浦肯野神经元及其轴突和树突界定,还有星状细胞、篮状细胞和高尔基细胞及其突起。在蚓部有一条狭窄的中线带,两侧各有三条较宽的带,每条带之间由无反应区隔开。尽管带区边界清晰,但带区之间的区域包含一些CSADCase免疫反应性轴突,但没有细胞体。七个蚓部带在前叶最易观察到。其他带存在于外侧半球。旁绒球和绒球包含大量强烈免疫反应性神经元,且带区难以辨别。蚓部的小叶X也富含牛磺酸能神经元。许多大、中型小脑深部核团和前庭核团也具有免疫反应性。这些观察结果表明,小脑神经元在化学性质上是异质的,但小脑皮质中具有相似化学特征的神经元被组织成矢状微带。这证实了我们早期的证据,即含有胃动素的浦肯野细胞以及同时含有胃动素和γ - 氨基丁酸的浦肯野细胞也排列在蚓部矢状微带中。中线蚓部带包含具有多种神经活性物质——牛磺酸、γ - 氨基丁酸和胃动素的浦肯野神经元。小脑皮质中的这种化学分区与来自脊髓、前庭、网状和橄榄核的带状传入神经支配之间的关系仍有待确定。