Proudfoot L, Schneider P, Ferguson M A, McConville M J
Department of Biochemistry, University of Dundee, Scotland, U.K.
Biochem J. 1995 May 15;308 ( Pt 1)(Pt 1):45-55. doi: 10.1042/bj3080045.
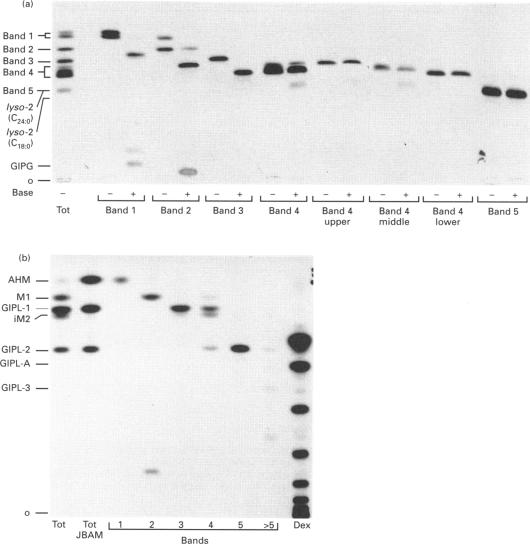
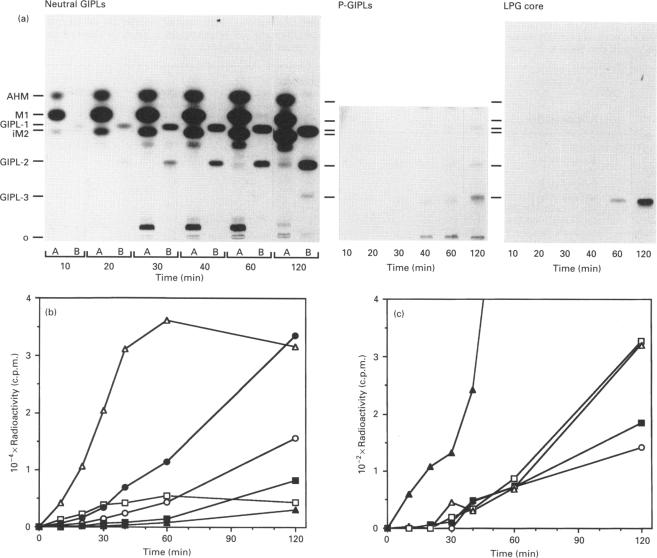
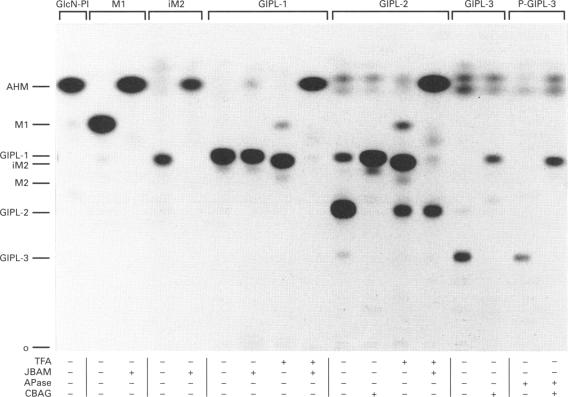
The major macromolecule on the surface of the protozoan parasite Leishmania major is a lipophosphoglycan (LPG) which contains a glycosylphosphatidylinositol glycolipid anchor. This parasite also synthesizes a complex family of abundant low-molecular-mass glycoinositolphospholipids (GIPLs) which are structurally related to the LPG anchor. In this study, L. major promastigotes were metabolically labelled with [3H]GlcN, and the kinetics of incorporation into free glycolipids and the LPG anchor followed to elucidate the pathway of GIPL biosynthesis and possible precursor-product relationships between the GIPLs and LPG. Labelled GIPLs were identified by TLC and by liquid chromatography of the released headgroups, before and after enzymic and chemical cleavage. On the basis of the measured specific radioactivities of the GIPLs, and their kinetics of radiolabelling, we suggest the pathway GlcN-PI-->Man1GlcN-PI (M1)-->Man2GlcN-PI (iM2)-->GalfMan2GlcN-PI (GIPL-1)-->Gal1GalfMan2GlcN-PI (GIPL-2)-->Gal2GalfMan2GlcN-PI (GIPL-3). All of the GIPLs were shown to contain alkylacylglycerol or lyso-alkylglycerol lipid moieties with the exception of the earliest intermediate, glucosaminylphosphatidylinositol (GlcN-PI), which contained both alkylacylglycerol and diacylglycerol. A significant proportion (approx. 50%) of GIPL-3 appeared to be selectively modified by the addition of a Glc-1-PO4 residue to one of the mannose residues (P-GIPL-3). On the basis of the specific radioactivity and kinetics of labelling of GIPL-3 and P-GIPL-3 we suggest that both of these low-abundance species are rapidly utilized as LPG precursors. The turnover of LPG and the GIPLs was also studied by [3H]Gal pulse-chase labelling and cell-surface labelling experiments. Whereas LPG was rapidly shed from the cell surface, consistent with previous studies, the GIPLs (both the total cellular and cell-surface pools) had a much slower turnover. These results suggest that the majority of the GIPLs do not act as LPG precursors and indicate that the cellular levels of these molecules is determined, at least in part, by the rate at which they are shed from the cell surface.
原生动物寄生虫硕大利什曼原虫表面的主要大分子是一种脂磷壁酸聚糖(LPG),它含有一个糖基磷脂酰肌醇糖脂锚定物。这种寄生虫还合成了一类复杂的丰富的低分子量糖基肌醇磷脂(GIPLs),它们在结构上与LPG锚定物相关。在本研究中,用[3H]葡糖胺对硕大利什曼原虫前鞭毛体进行代谢标记,并追踪其掺入游离糖脂和LPG锚定物的动力学,以阐明GIPL生物合成途径以及GIPLs与LPG之间可能的前体-产物关系。在酶解和化学裂解前后,通过薄层层析(TLC)和释放的头部基团的液相色谱法鉴定标记的GIPLs。根据测得的GIPLs的比放射性及其放射性标记动力学,我们提出了如下途径:葡糖胺-磷脂酰肌醇(GlcN-PI)→甘露糖1葡糖胺-磷脂酰肌醇(M1)→甘露糖2葡糖胺-磷脂酰肌醇(iM2)→半乳糖基甘露糖2葡糖胺-磷脂酰肌醇(GIPL-1)→半乳糖1半乳糖基甘露糖2葡糖胺-磷脂酰肌醇(GIPL-2)→半乳糖2半乳糖基甘露糖2葡糖胺-磷脂酰肌醇(GIPL-3)。除了最早的中间体葡糖胺基磷脂酰肌醇(GlcN-PI)同时含有烷基酰基甘油和二酰基甘油外,所有的GIPLs都显示含有烷基酰基甘油或溶血烷基酰基甘油脂质部分。相当一部分(约50%)的GIPL-3似乎通过在其中一个甘露糖残基上添加一个葡糖-1-磷酸残基而被选择性修饰(P-GIPL-3)。根据GIPL-3和P-GIPL-3的比放射性和标记动力学,我们认为这两种低丰度物质都迅速被用作LPG前体。还通过[3H]半乳糖脉冲追踪标记和细胞表面标记实验研究了LPG和GIPLs的周转情况。与先前的研究一致,LPG迅速从细胞表面脱落,而GIPLs(细胞内和细胞表面池中的总量)的周转则慢得多。这些结果表明,大多数GIPLs不作为LPG前体,并且表明这些分子的细胞水平至少部分由它们从细胞表面脱落的速率决定。