Cullinane C, Cutts S M, van Rosmalen A, Phillips D R
Department of Biochemistry, La Trobe University, Bundoora, Victoria, Australia.
Nucleic Acids Res. 1994 Jun 25;22(12):2296-303. doi: 10.1093/nar/22.12.2296.
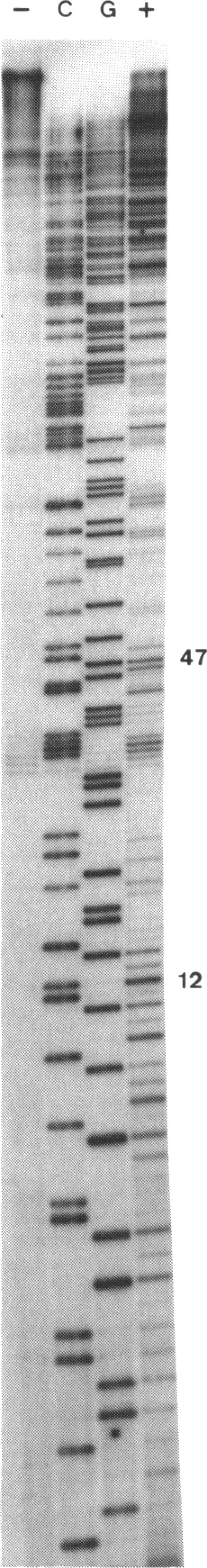
Adriamycin is known to induce the formation of adducts with DNA when reacted under in vitro transcription conditions. The factors affecting the extent of adduct formation were examined in order to establish the critical components and optimal conditions required for the reaction, and to gain insight into the nature of the DNA-adduct complex. There was a strong dependence on reaction temperature (with a 40-fold increase of adducts at 40-50 degrees C compared to 10 degrees C), pH (maximum adducts at pH 7), but little dependence on the oxygen level. There was an absolute requirement for a reducing agent, with adducts detected with DTT, beta-mercaptoethanol and glutathione, maximal adducts were formed at high levels of DTT (5-10 mM). Adducts were also formed with a xanthine oxidase/NADH reducing system, with increasing amounts of adducts detected with increasing NADH; no adducts were detected in the absence of either the enzyme or NADH. Of fourteen derivatives studied, only four yielded a similar extent of adduct formation as adriamycin; there was no absolute requirement for a carbonyl at C13 or hydroxyl at C14. Adducts were also observed with ssDNA but required a longer reaction time compared to dsDNA. The sequence specificity of adduct formation with ssDNA was examined using a primer-extension assay; almost all adducts were associated with a guanine residue. Overall, the results are consistent with a two-step reaction mechanism involving reductive activation of adriamycin, with the activated species then reacting with the guanine residues of either dsDNA or ssDNA.
已知阿霉素在体外转录条件下反应时会诱导与DNA形成加合物。为确定反应所需的关键成分和最佳条件,并深入了解DNA - 加合物复合物的性质,研究了影响加合物形成程度的因素。加合物形成程度强烈依赖于反应温度(40 - 50摄氏度时的加合物比10摄氏度时增加40倍)、pH值(pH 7时加合物最多),但对氧气水平依赖性较小。反应绝对需要还原剂,用二硫苏糖醇(DTT)、β - 巯基乙醇和谷胱甘肽可检测到加合物,在高浓度DTT(5 - 10 mM)时形成的加合物最多。用黄嘌呤氧化酶/NADH还原系统也能形成加合物,随着NADH增加检测到的加合物量增多;在没有酶或NADH时未检测到加合物。在所研究的14种衍生物中,只有4种产生的加合物形成程度与阿霉素相似;对C13处的羰基或C14处的羟基没有绝对要求。与单链DNA(ssDNA)也能形成加合物,但与双链DNA(dsDNA)相比需要更长的反应时间。使用引物延伸试验检测了与ssDNA形成加合物的序列特异性;几乎所有加合物都与鸟嘌呤残基相关。总体而言,结果与涉及阿霉素还原活化的两步反应机制一致,活化后的物种随后与dsDNA或ssDNA的鸟嘌呤残基反应。