Myrset A H, Bostad A, Jamin N, Lirsac P N, Toma F, Gabrielsen O S
Department of Biochemistry, University of Oslo, Norway.
EMBO J. 1993 Dec;12(12):4625-33. doi: 10.1002/j.1460-2075.1993.tb06151.x.
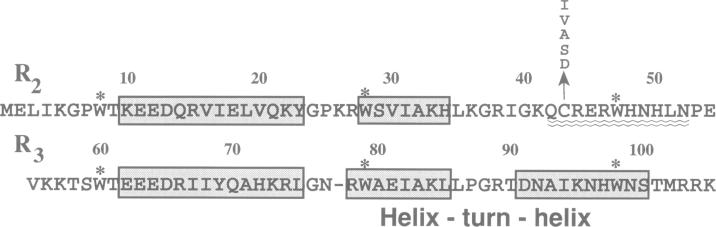
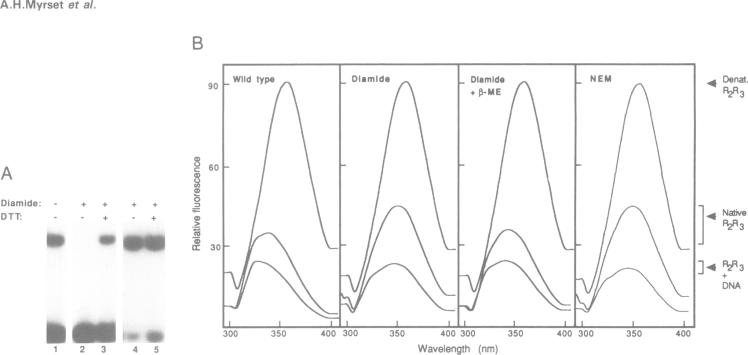
The DNA-binding domain of the oncoprotein Myb comprises three imperfect repeats, R1, R2 and R3. Only R2 and R3 are required for sequence-specific DNA-binding. Both are assumed to contain helix-turn-helix (HTH)-related motifs, but multidimensional heteronuclear NMR spectroscopy revealed a disordered structure in R2 where the second HTH helix was predicted [Jamin et al. (1993) Eur. J. Biochem., 216, 147-154]. We propose that the disordered region folds into a 'recognition' helix and generates a full HTH-related motif upon binding to DNA. This would move Cys43 into the hydrophobic core of R2. We observed that Cys43 was accessible to N-ethylmaleimide alkylation in the free protein, but inaccessible in the DNA complex. Mutant proteins with charged (C43D) or polar (C43S) side chains in position 43 bound DNA with reduced affinity, while hydrophobic replacements (C43A, C43V and C43I) gave unaltered or improved DNA-binding. Specific DNA-binding enhanced protease resistance dramatically. Fluorescence emission spectra and quenching experiments supported a DNA-induced conformational change. Moreover, reversible oxidation of Cys43 had an effect similar to the inactivating C43D mutation. The highly oxidizable Cys43 could function as a molecular sensor for a redox regulatory mechanism turning specific DNA-binding on or off by controlling the DNA-induced conformational change in R2.
癌蛋白Myb的DNA结合结构域由三个不完全重复序列R1、R2和R3组成。序列特异性DNA结合仅需要R2和R3。两者都被认为包含与螺旋-转角-螺旋(HTH)相关的基序,但多维异核核磁共振光谱显示R2中存在无序结构,其中预测了第二个HTH螺旋[贾明等人(1993年)《欧洲生物化学杂志》,216卷,147 - 154页]。我们提出,无序区域折叠成一个“识别”螺旋,并在与DNA结合时形成一个完整的与HTH相关的基序。这将使Cys43进入R2的疏水核心。我们观察到,在游离蛋白中Cys43可被N - 乙基马来酰亚胺烷基化,但在DNA复合物中则不可。在第43位带有带电(C43D)或极性(C43S)侧链的突变蛋白与DNA结合的亲和力降低,而疏水替代(C43A、C43V和C43I)则使DNA结合不变或增强。特异性DNA结合显著增强了蛋白酶抗性。荧光发射光谱和猝灭实验支持了DNA诱导的构象变化。此外,Cys43的可逆氧化具有与失活的C43D突变类似的效果。高度可氧化的Cys43可作为一种分子传感器,用于通过控制R2中DNA诱导的构象变化来开启或关闭特异性DNA结合的氧化还原调节机制。