Stone J C, Colleton M, Bottorff D
Department of Biochemistry, University of Alberta, Edmonton, Canada.
Mol Cell Biol. 1993 Dec;13(12):7311-20. doi: 10.1128/mcb.13.12.7311-7320.1993.
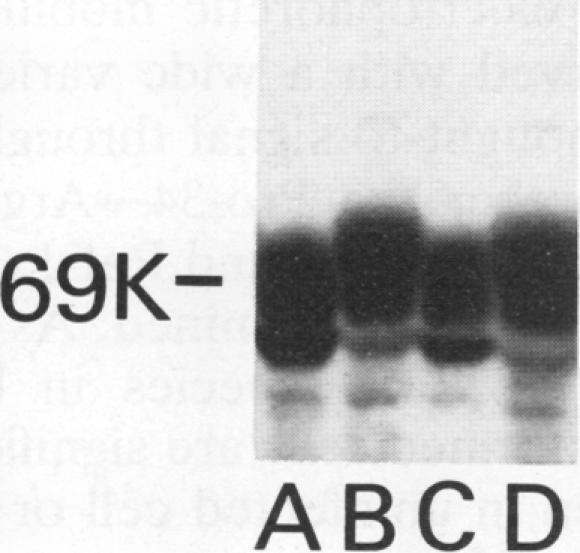
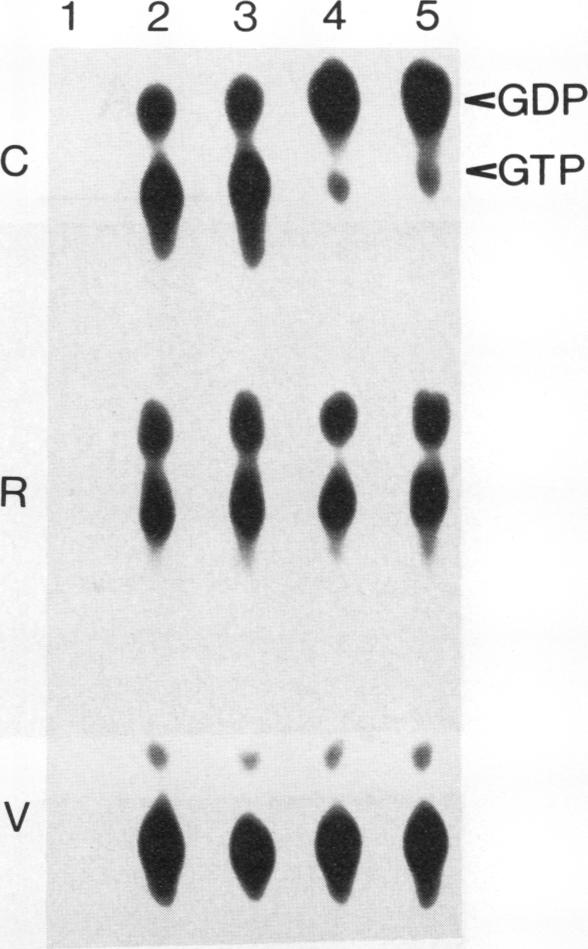
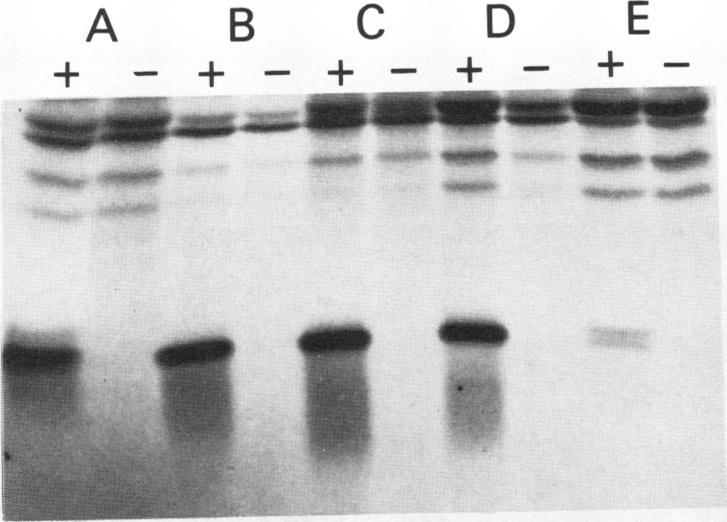
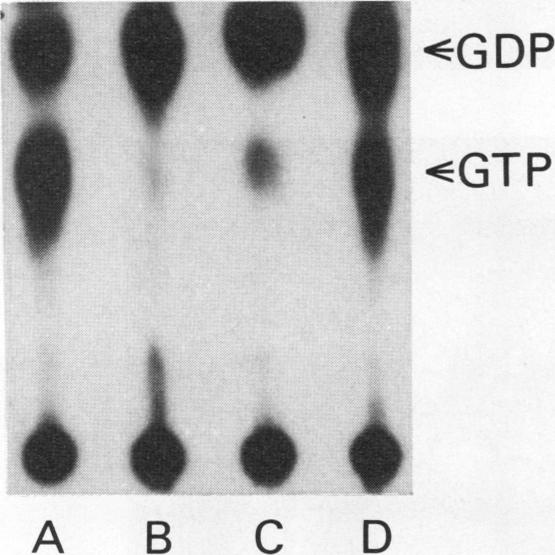
The GTPase activity of p21ras is stimulated by GTPase-activating proteins (GAPs) such as p120GAP and the product of the neurofibromatosis 1 gene, which may negatively regulate p21 function. GAPs are also proposed effectors of ras. We have sought activating substitutions in c-H-ras in the region encoding the effector domain, on the rationale that such mutations would dissociate effector function from negative regulation by GAP. One such activating mutation, Pro-34-->Arg, encodes protein that is substantially bound to GTP in vivo. In vitro, this protein is not stimulated by GAPs, and its binding to p120GAP is grossly impaired. The results support the idea that the p21 structural requirements for effector function and GAP interaction are quite different and suggest that some molecule(s) other than p120GAP serves as the ras effector. In contrast to the results obtained with p120GAP, the Pro-34-->Arg p21 species is effectively coupled to the raf-1 product, as judged from electrophoretic mobility shifts of the Raf-1 phosphoprotein.
p21ras的GTP酶活性受GTP酶激活蛋白(GAPs)如p120GAP和神经纤维瘤病1基因产物的刺激,这些蛋白可能对p21功能起负调节作用。GAPs也被认为是ras的效应器。我们在编码效应器结构域的区域寻找c-H-ras中的激活替代,基于这样的理论,即此类突变会使效应器功能与GAP的负调节分离。一种这样的激活突变,Pro-34→Arg,编码的蛋白在体内基本上与GTP结合。在体外,该蛋白不受GAPs刺激,并且其与p120GAP的结合严重受损。这些结果支持这样的观点,即效应器功能和GAP相互作用的p21结构要求截然不同,并表明除p120GAP之外的某些分子作为ras效应器。与用p120GAP获得的结果相反,根据Raf-1磷酸化蛋白的电泳迁移率变化判断,Pro-34→Arg p21物种有效地与raf-1产物偶联。