Tran P T, Walker R A, Salmon E D
Biology Department, University of North Carolina, Chapel Hill, North Carolina 27599-3280, USA.
J Cell Biol. 1997 Jul 14;138(1):105-17. doi: 10.1083/jcb.138.1.105.
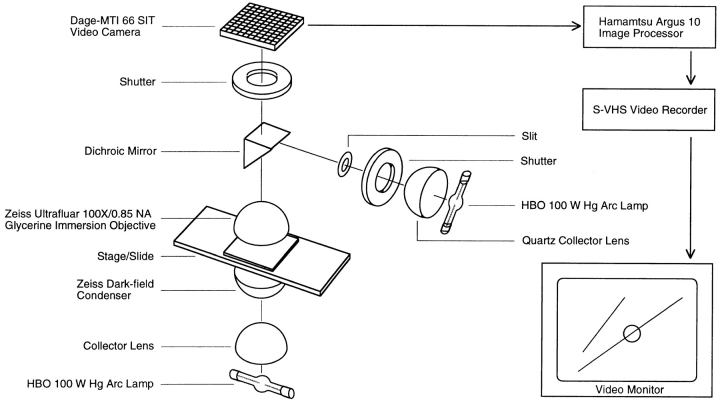
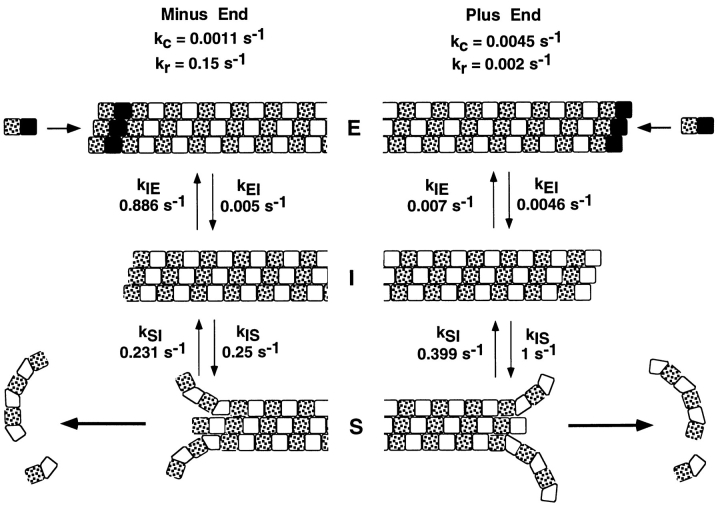
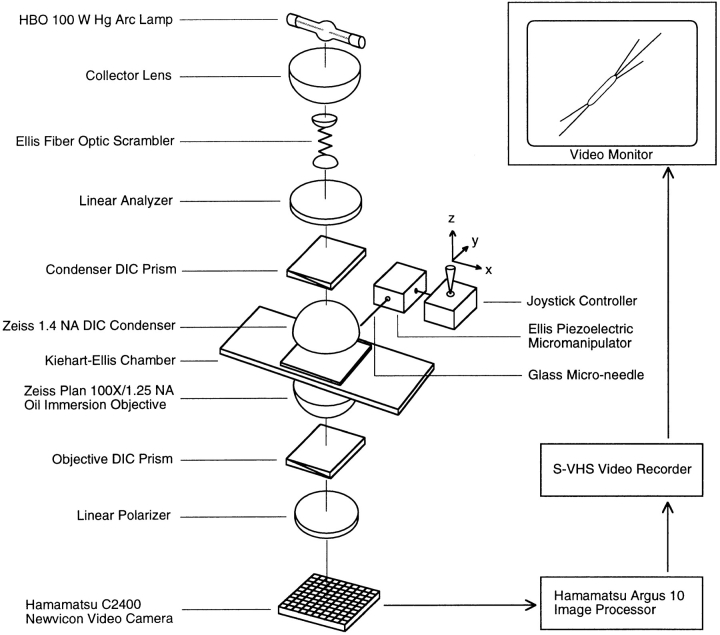
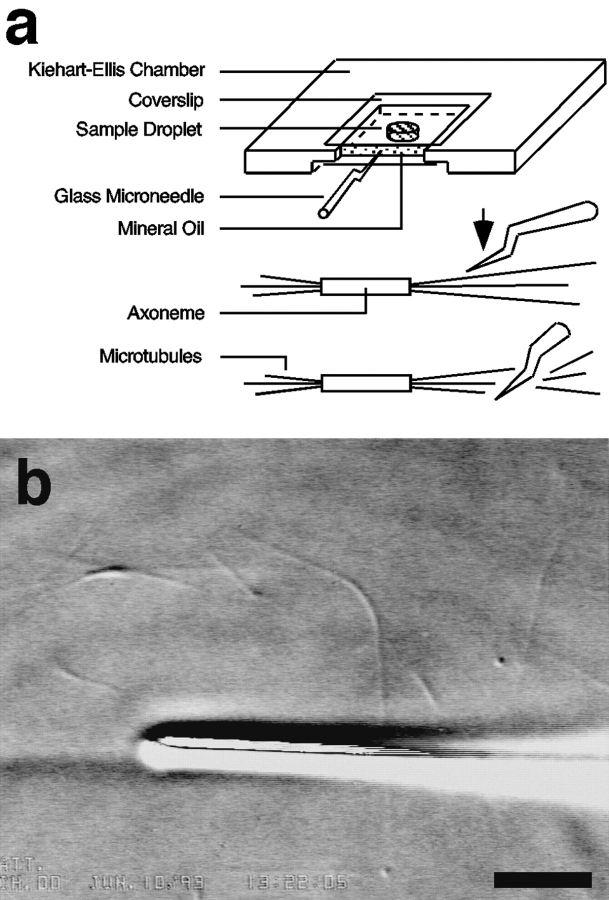
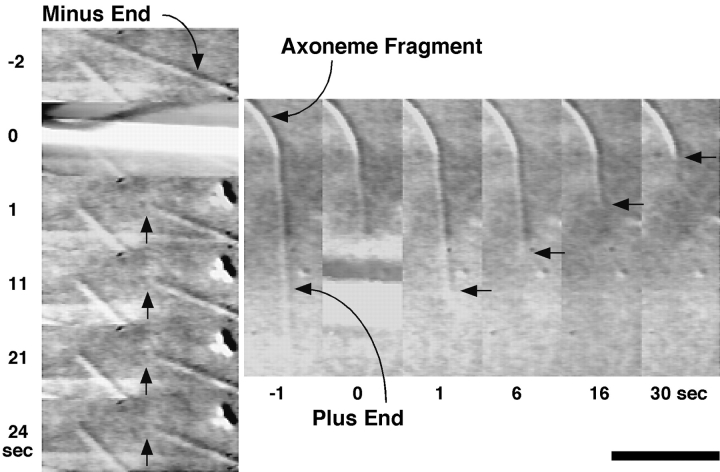
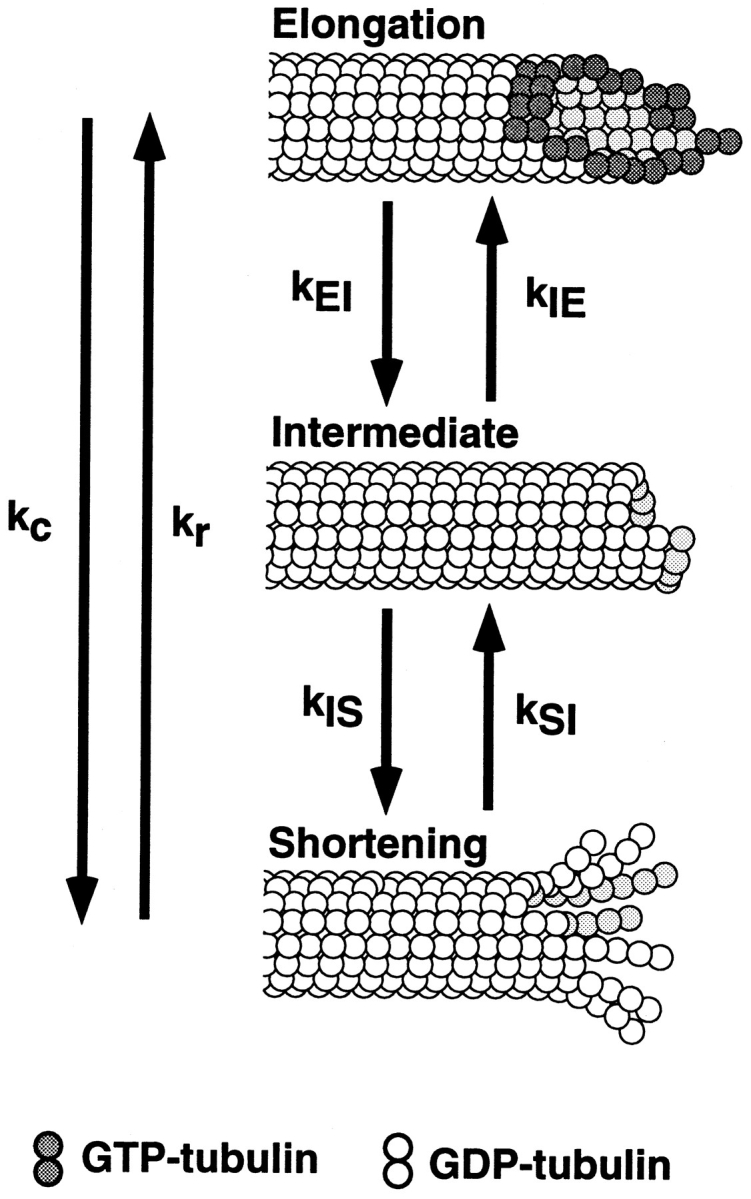
The current two-state GTP cap model of microtubule dynamic instability proposes that a terminal crown of GTP-tubulin stabilizes the microtubule lattice and promotes elongation while loss of this GTP-tubulin cap converts the microtubule end to shortening. However, when this model was directly tested by using a UV microbeam to sever axoneme-nucleated microtubules and thereby remove the microtubule's GTP cap, severed plus ends rapidly shortened, but severed minus ends immediately resumed elongation (Walker, R.A., S. Inoué, and E.D. Salmon. 1989. J. Cell Biol. 108: 931-937). To determine if these previous results were dependent on the use of axonemes as seeds or were due to UV damage, or if they instead indicate an intermediate state in cap dynamics, we performed UV cutting of self-assembled microtubules and mechanical cutting of axoneme-nucleated microtubules. These independent methods yielded results consistent with the original work: a significant percentage of severed minus ends are stable after cutting. In additional experiments, we found that the stability of both severed plus and minus ends could be increased by increasing the free tubulin concentration, the solution GTP concentration, or by assembling microtubules with guanylyl-(alpha,beta)-methylene-diphosphonate (GMPCPP). Our results show that stability of severed ends, particularly minus ends, is not an artifact, but instead reveals the existence of a metastable kinetic intermediate state between the elongation and shortening states of dynamic instability. The kinetic properties of this intermediate state differ between plus and minus ends. We propose a three-state conformational cap model of dynamic instability, which has three structural states and four transition rate constants, and which uses the asymmetry of the tubulin heterodimer to explain many of the differences in dynamic instability at plus and minus ends.
目前关于微管动态不稳定性的两态GTP帽模型提出,GTP - 微管蛋白的末端冠稳定微管晶格并促进伸长,而这种GTP - 微管蛋白帽的丢失会使微管末端转变为缩短状态。然而,当使用紫外线微束切断轴丝成核的微管从而去除微管的GTP帽来直接测试该模型时,切断的正端迅速缩短,但切断的负端立即恢复伸长(Walker, R.A., S. Inoué, and E.D. Salmon. 1989. J. Cell Biol. 108: 931 - 937)。为了确定这些先前的结果是依赖于使用轴丝作为种子还是由于紫外线损伤,或者它们是否反而表明帽动态中的一种中间状态,我们对自组装微管进行了紫外线切割,并对轴丝成核的微管进行了机械切割。这些独立的方法得出的结果与原始工作一致:相当比例的切断负端在切割后是稳定的。在额外的实验中,我们发现通过增加游离微管蛋白浓度、溶液GTP浓度或用鸟苷酰 -(α,β)-亚甲基二磷酸酯(GMPCPP)组装微管,可以提高切断的正端和负端的稳定性。我们的结果表明,切断末端的稳定性,特别是负端,不是人为现象,而是揭示了动态不稳定性的伸长和缩短状态之间存在亚稳态动力学中间状态。这种中间状态的动力学特性在正端和负端之间有所不同。我们提出了一个动态不稳定性的三态构象帽模型,该模型有三个结构状态和四个转变速率常数,并利用微管蛋白异二聚体的不对称性来解释正端和负端在动态不稳定性方面的许多差异。