Kaftan E J, Ehrlich B E, Watras J
Department of Physiology, University of Connecticut, Farmington, Connecticut 06030-3505, USA.
J Gen Physiol. 1997 Nov;110(5):529-38. doi: 10.1085/jgp.110.5.529.
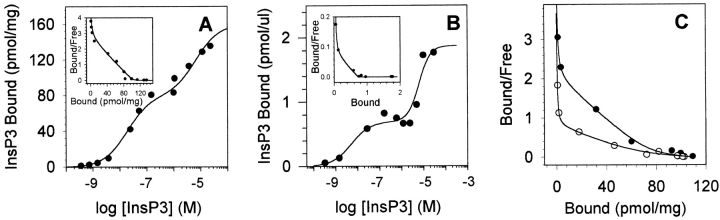
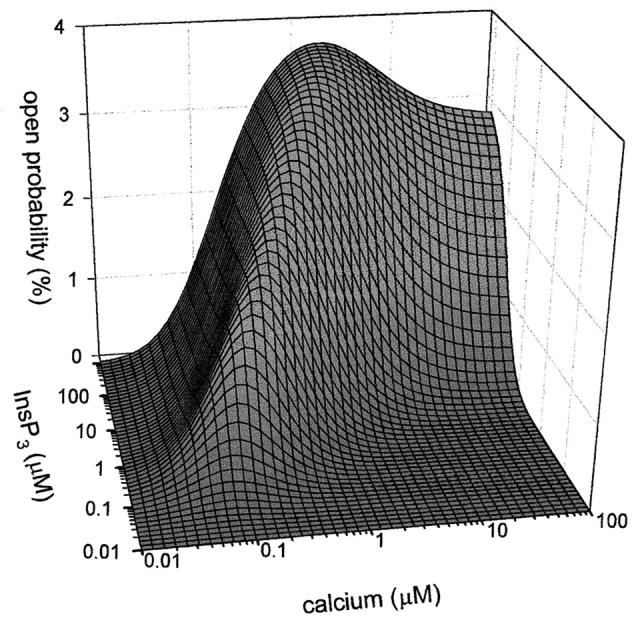
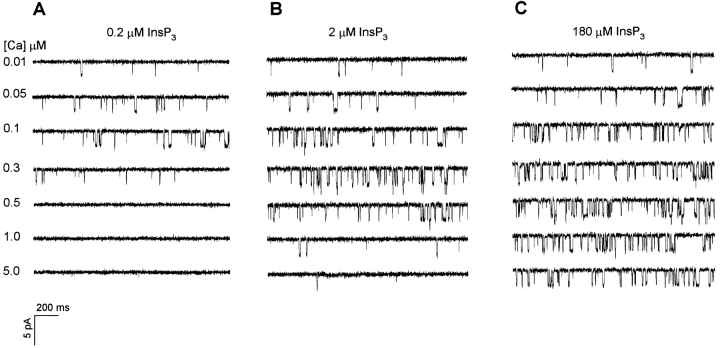
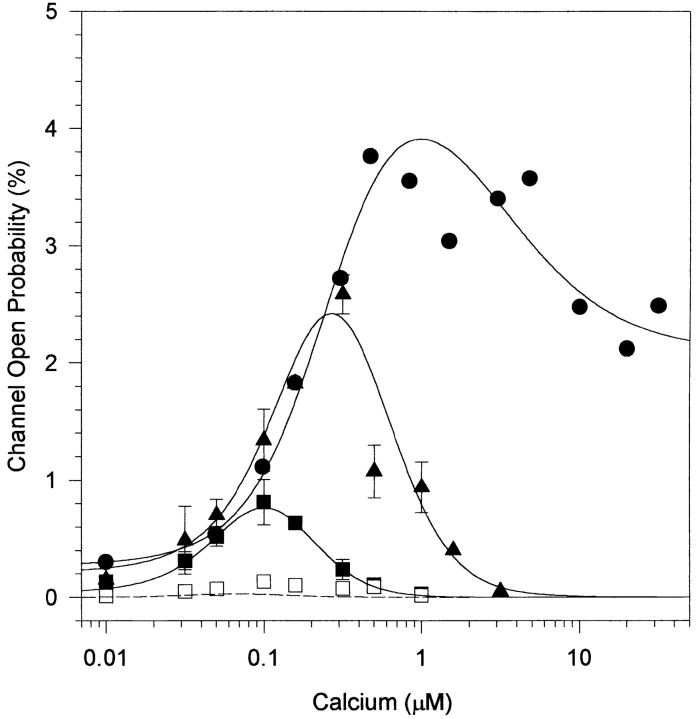
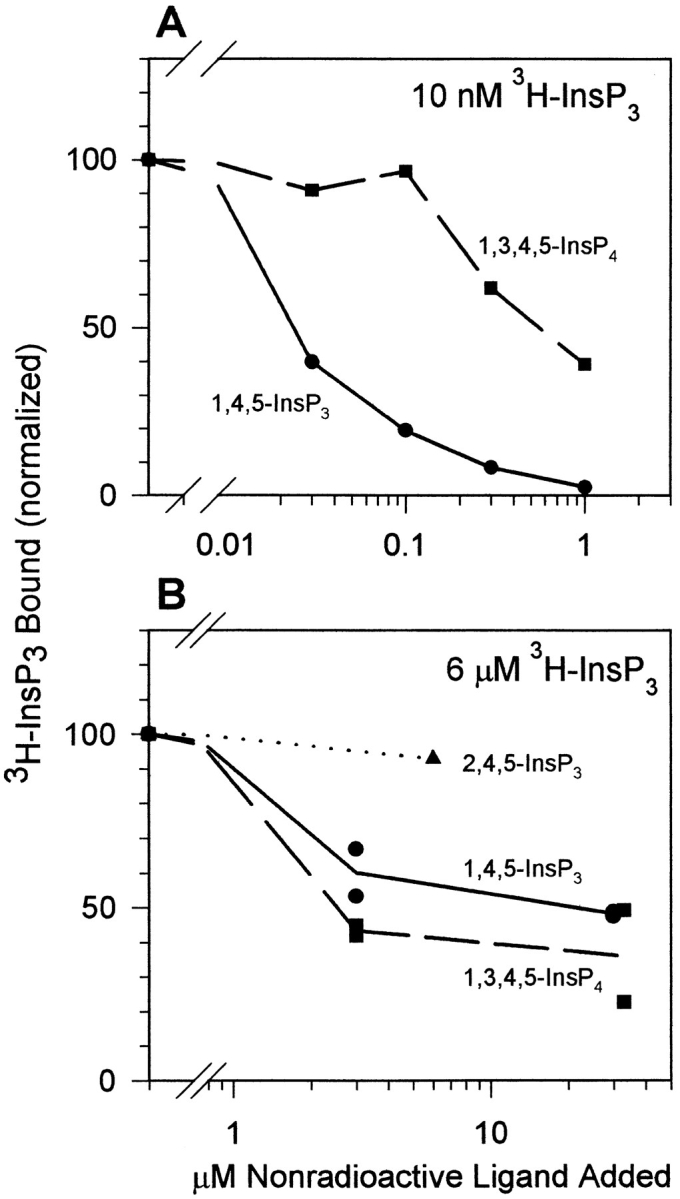
The inositol 1,4,5-trisphosphate (InsP3)-gated Ca channel in cerebellum is tightly regulated by Ca (Bezprozvanny, I., J. Watras, and B.E. Ehrlich. 1991. Nature (Lond.). 351:751-754; Finch, E.A., T. J. Turner, and S.M. Goldin. 1991. Science (Wash. DC). 252:443-446; Hannaert-Merah, Z., J.F. Coquil, L. Combettes, M. Claret, J.P. Mauger, and P. Champeil. 1994. J. Biol. Chem. 269:29642-29649; Iino, M. 1990. J. Gen. Physiol. 95:1103-1122; Marshall, I., and C. Taylor. 1994. Biochem. J. 301:591-598). In previous single channel studies, the Ca dependence of channel activity, monitored at 2 microM InsP3, was described by a bell-shaped curve (Bezprozvanny, I., J. Watras, and B.E. Ehrlich. 1991. Nature (Lond.). 351:751-754). We report here that, when we used lower InsP3 concentrations, the peak of the Ca-dependence curve shifted to lower Ca concentrations. Unexpectedly, when we used high InsP3 concentrations, channel activity persisted at Ca concentrations as high as 30 microM. To explore this unexpected response of the channel, we measured InsP3 binding over a broad range of InsP3 concentrations. We found the well-characterized high affinity InsP3 binding sites (with Kd < 1 and 50 nM) (Maeda, N., M. Niinobe, and K. Mikoshiba. 1990. EMBO (Eur. Mol. Biol. Organ.) J. 9:61-67; Mignery, G., T.C. Sudhof, K. Takei, and P. De Camilli. 1989. Nature (Lond.). 342:192-195; Ross, C.A., J. Meldolesi, T.A. Milner, T. Satoh, S. Supattapone, and S.H. Snyder. 1989. Nature (Lond.). 339:468-470) and a low affinity InsP3 binding site (Kd = 10 microM). Using these InsP3 binding sites, we developed a new model that accounts for the shift in the Ca-dependence curve at low InsP3 levels and the maintained channel activity at high Ca and InsP3 levels. The observed Ca dependence of the InsP3-gated Ca channel allows the cell to abbreviate the rise of intracellular Ca in the presence of low levels of InsP3, but also provides a means of maintaining high intracellular Ca during periods of prolonged stimulation.
小脑肌醇 1,4,5 - 三磷酸(InsP3)门控钙通道受钙离子严格调控(贝兹普罗兹瓦尼,I.,J. 瓦特拉,和 B.E. 埃利希。1991 年。《自然》(伦敦)。351:751 - 754;芬奇,E.A.,T.J. 特纳,和 S.M. 戈尔丁。1991 年。《科学》(华盛顿特区)。252:443 - 446;汉纳尔特 - 梅拉,Z.,J.F. 科基尔,L. 孔贝茨,M. 克莱雷,J.P. 莫热,和 P. 尚佩伊。1994 年。《生物化学杂志》。269:29642 - 29649;饭野,M. 1990 年。《普通生理学杂志》。95:1103 - 1122;马歇尔,I.,和 C. 泰勒。1994 年。《生物化学杂志》。301:591 - 598)。在先前的单通道研究中,在 2 微摩尔 InsP3 浓度下监测到的通道活性对钙离子的依赖性由一条钟形曲线描述(贝兹普罗兹瓦尼,I.,J. 瓦特拉,和 B.E. 埃利希。1991 年。《自然》(伦敦)。351:751 - 754)。我们在此报告,当使用较低的 InsP3 浓度时,钙离子依赖性曲线的峰值向较低的钙离子浓度偏移。出乎意料的是,当使用高 InsP3 浓度时,通道活性在高达 30 微摩尔的钙离子浓度下仍持续存在。为探究通道的这种意外反应,我们在广泛的 InsP3 浓度范围内测量了 InsP3 结合情况。我们发现了特征明确的高亲和力 InsP3 结合位点(解离常数 Kd < 1 和 50 纳摩尔)(前田,N.,M. 新之部,和 K. 三木志保。1990 年。《欧洲分子生物学组织杂志》。9:61 - 67;米涅里,G.,T.C. 苏德霍夫,K. 武井,和 P. 德卡米利。1989 年。《自然》(伦敦)。342:192 - 195;罗斯,C.A.,J. 梅尔多莱西,T.A. 米尔纳,T. 佐藤,S. 苏帕塔蓬内,和 S.H. 斯奈德。1989 年。《自然》(伦敦)。339:468 - 470)以及一个低亲和力 InsP3 结合位点(Kd = 10 微摩尔)。利用这些 InsP3 结合位点,我们建立了一个新模型,该模型解释了在低 InsP3 水平下钙离子依赖性曲线的偏移以及在高钙离子和 InsP3 水平下通道活性的维持。所观察到的 InsP3 门控钙通道对钙离子的依赖性使细胞在低水平 InsP3 存在时能够缩短细胞内钙离子的升高,但也提供了一种在长时间刺激期间维持高细胞内钙离子水平的方式。