Nicchitta C V, Zheng T
Department of Cell Biology, Duke University Medical Center, Durham, North Carolina 27710, USA.
J Cell Biol. 1997 Dec 29;139(7):1697-708. doi: 10.1083/jcb.139.7.1697.
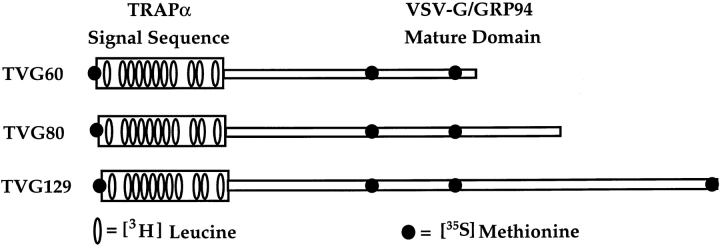
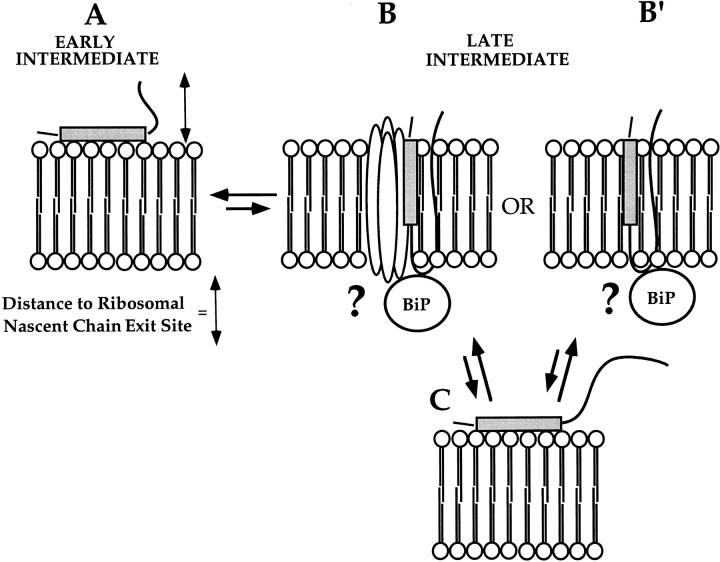
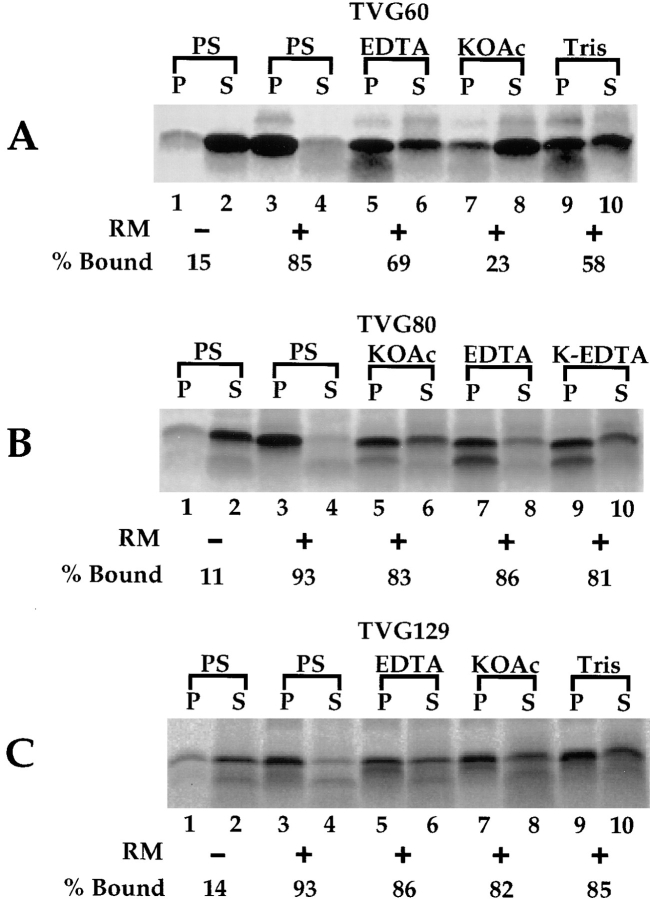
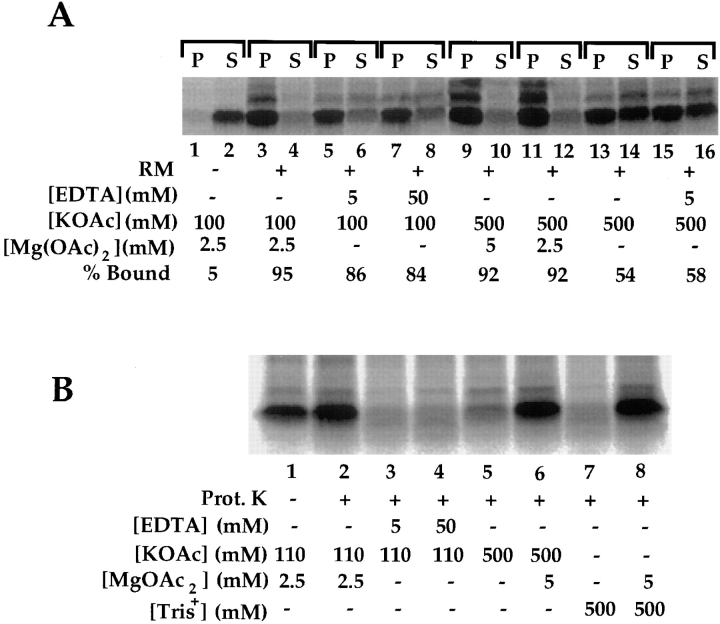
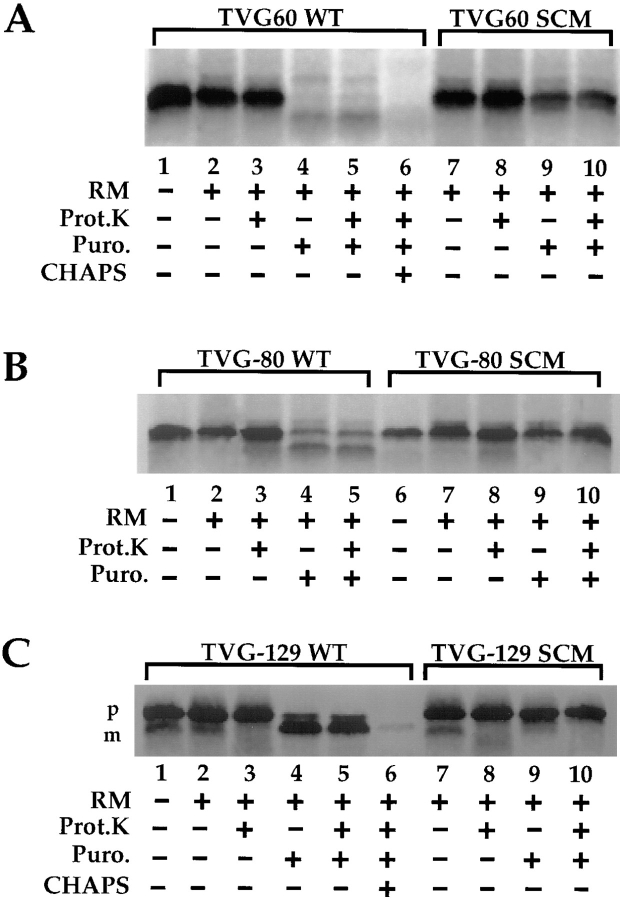
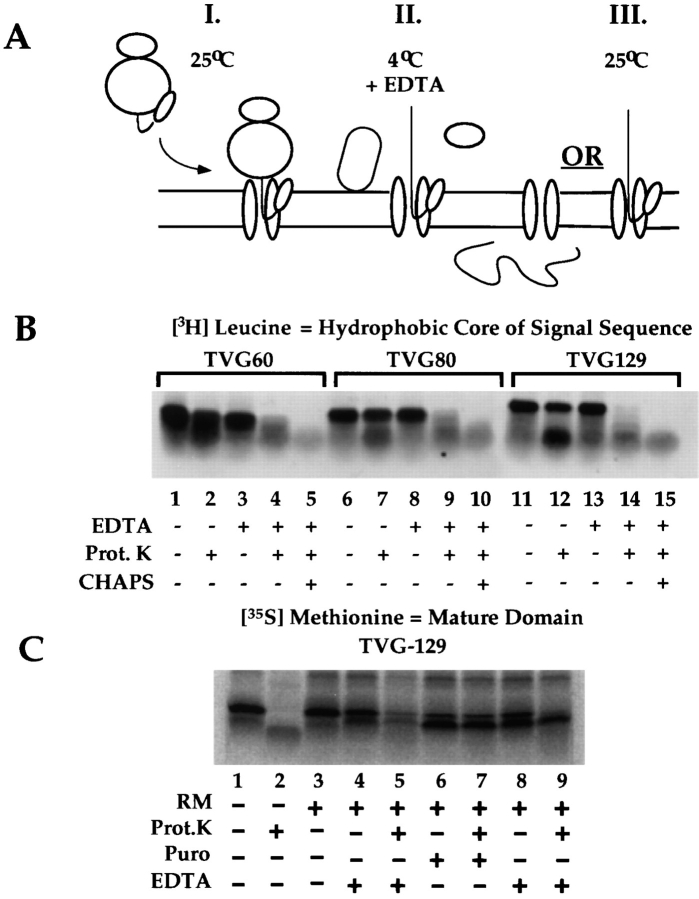
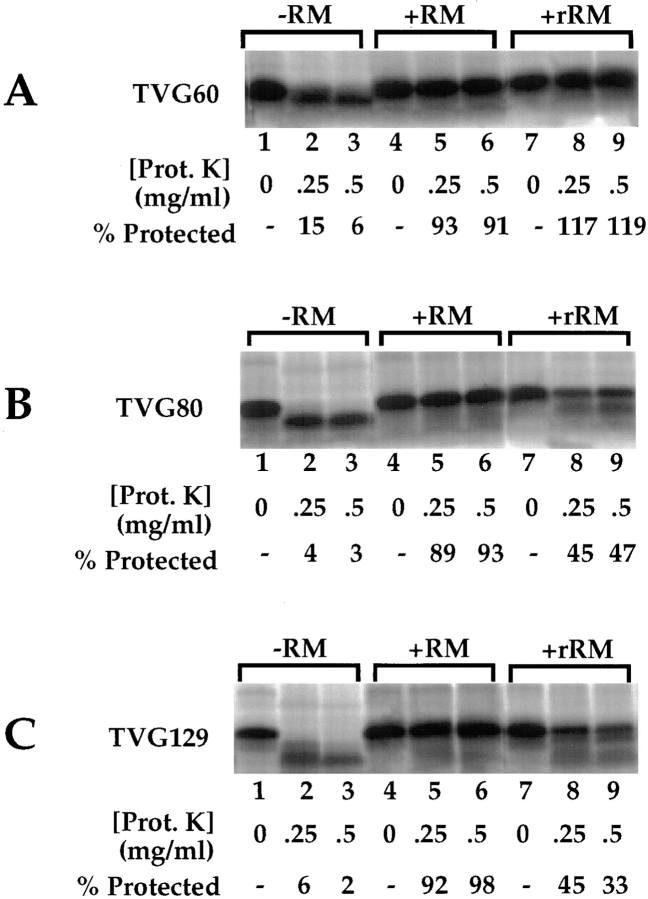
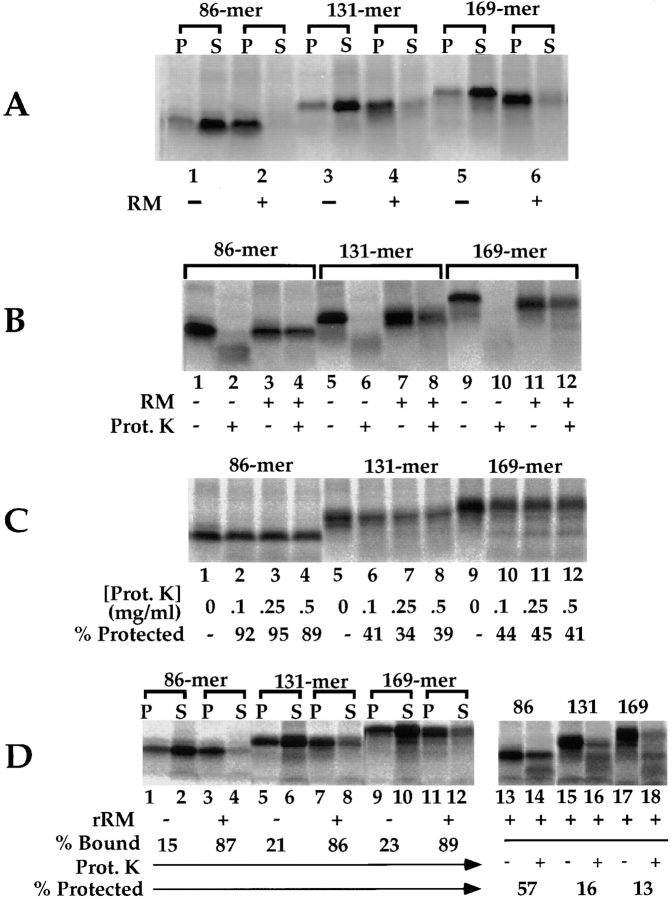
A series of fusion protein constructs were designed to investigate the contribution of secretory nascent chains to regulation of the ribosome-membrane junction in the mammalian endoplasmic reticulum. As a component of these studies, the membrane topology of the signal sequence was determined at stages of protein translocation immediately after targeting and before signal sequence cleavage. Truncated translation products were used to delimit the analysis to defined stages of translocation. In a study of secretory protein precursors, formation of a protease-resistant ribosome-membrane junction, currently thought to define the pathway of the translocating nascent chain, was observed to be precursor- and stage-dependent. Analysis of the binding of early intermediates indicated that the nascent chain was bound to the membrane independent of the ribosome, and that the binding was predominately electrostatic. The membrane topology of the signal sequence was determined as a function of the stage of translocation, and was found to be identical for all assayed intermediates. Unexpectedly, the hydrophobic core of the signal sequence was observed to be accessible to the cytosolic face of the membrane at stages of translocation immediately after targeting as well as stages before signal sequence cleavage. Removal of the ribosome from bound intermediates did not disrupt subsequent translocation, suggesting that the active state of the protein-conducting channel is maintained in the absence of the bound ribosome. A model describing a potential mode of regulation of the ribosome-membrane junction by the nascent chain is presented.
设计了一系列融合蛋白构建体,以研究分泌新生链对哺乳动物内质网中核糖体 - 膜结合调节的作用。作为这些研究的一部分,在靶向之后且信号序列切割之前的蛋白质转运阶段确定信号序列的膜拓扑结构。使用截短的翻译产物将分析限定在转运的特定阶段。在一项分泌蛋白前体的研究中,观察到目前认为可定义转运新生链途径的蛋白酶抗性核糖体 - 膜结合的形成是前体和阶段依赖性的。对早期中间体结合的分析表明,新生链与膜的结合独立于核糖体,并且这种结合主要是静电作用。信号序列的膜拓扑结构是根据转运阶段确定的,并且发现所有测定的中间体都是相同的。出乎意料的是,在靶向之后以及信号序列切割之前的转运阶段,信号序列的疏水核心可被膜的胞质面接触。从结合的中间体中去除核糖体不会破坏随后的转运,这表明在没有结合核糖体的情况下,蛋白质传导通道的活性状态得以维持。提出了一个描述新生链对核糖体 - 膜结合潜在调节模式的模型。