Bailly M, Macaluso F, Cammer M, Chan A, Segall J E, Condeelis J S
Department of Anatomy and Structural Biology, Albert Einstein College of Medicine, Bronx, New York 10461, USA.
J Cell Biol. 1999 Apr 19;145(2):331-45. doi: 10.1083/jcb.145.2.331.
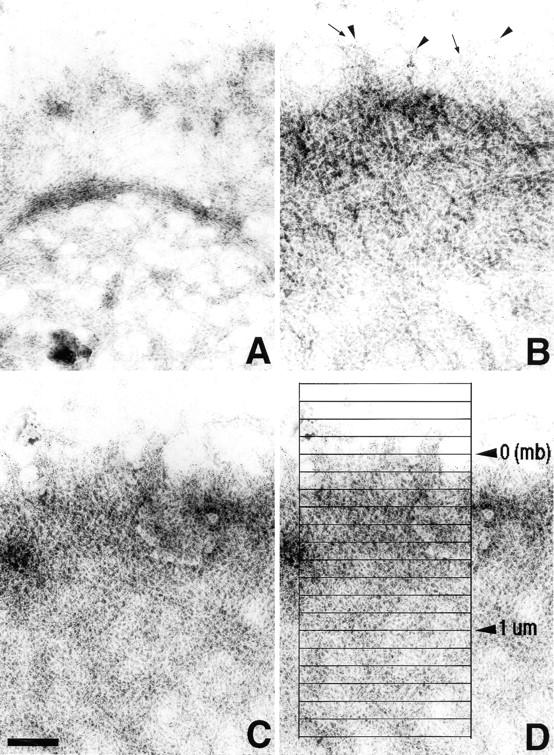
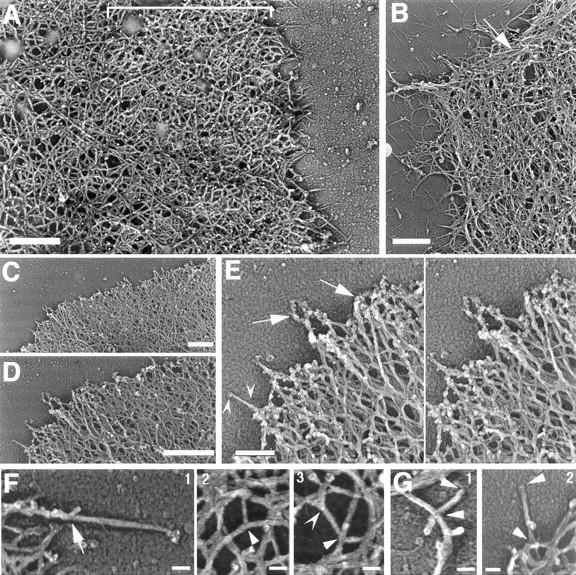
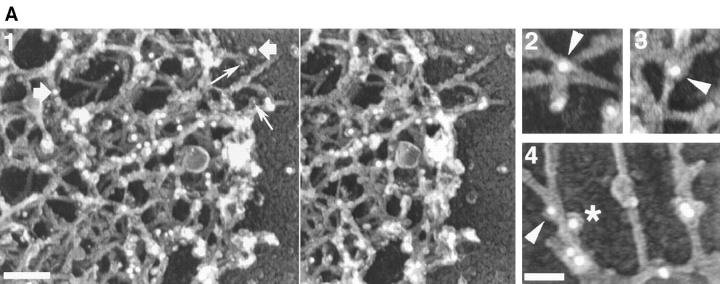
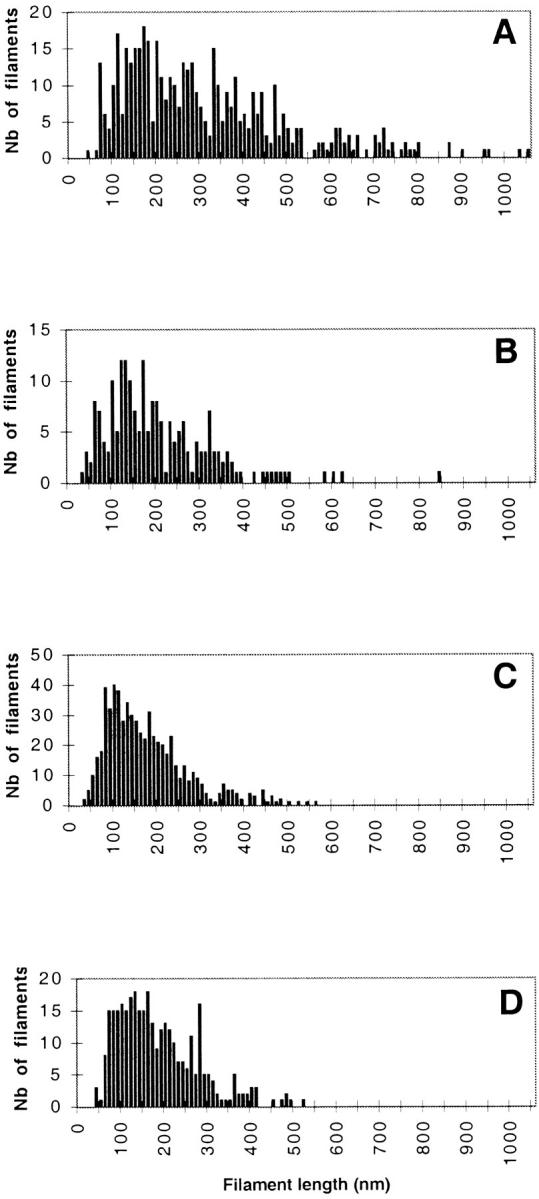
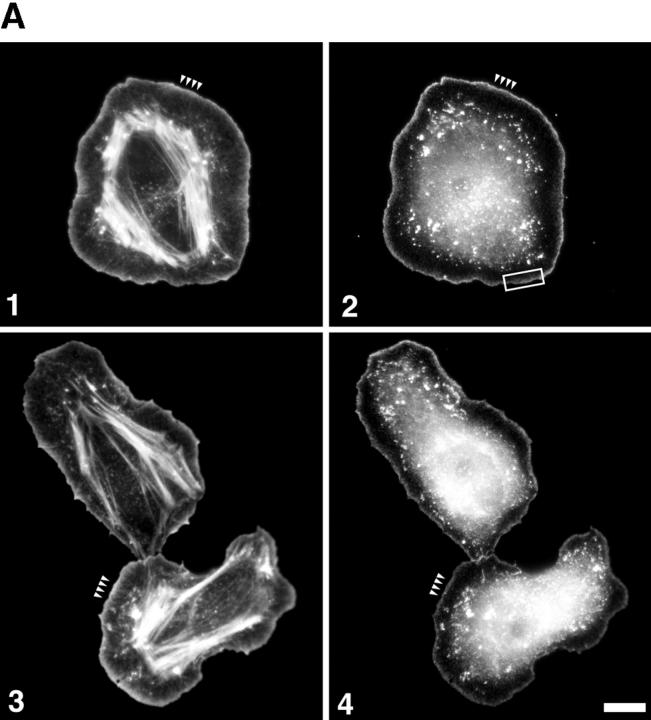
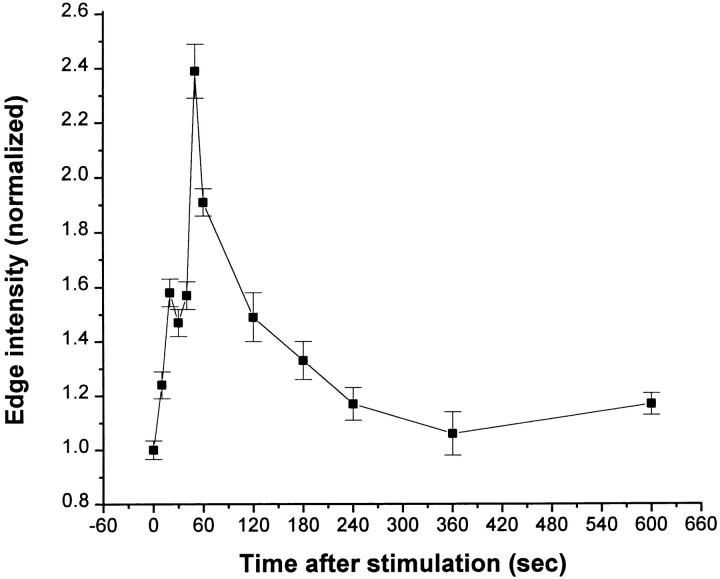
Using both light and high resolution electron microscopy, we analyzed the spatial and temporal relationships between the Arp2/3 complex and the nucleation activity that is required for lamellipod extension in mammary carcinoma cells after epidermal growth factor stimulation. A rapid two- to fourfold increase in filament barbed end number occurs transiently after stimulation and remains confined almost exclusively to the extreme outer edge of the extending lamellipod (within 100-200 nm of the plasma membrane). This is accompanied by an increase in filament density at the leading edge and a general decrease in filament length, with a specific loss of long filaments. Concomitantly, the Arp2/3 complex is recruited with a 1.5-fold increase throughout the entire cortical filament network extending 1-1.5 microm in depth from the membrane at the leading edge. The recruitment of the Arp2/3 complex at the membrane of the extending lamellipod indicates that Arp2/3 may be involved in initial generation of growing filaments. However, only a small subset of the complex present in the cortical network colocalizes near free barbed ends. This suggests that the 100-200-nm submembraneous compartment at the leading edge of the extending lamellipod constitutes a special biochemical microenvironment that favors the generation and maintenance of free barbed ends, possibly through the locally active Arp2/3 complex, severing or decreasing the on-rate of capping protein. Our results are inconsistent with the hypothesis suggesting uncapping is the dominant mechanism responsible for the generation of nucleation activity. However, they support the hypothesis of an Arp2/3-mediated capture of actin oligomers that formed close to the membrane by other mechanisms such as severing. They also support pointed-end capping by the Arp2/3 complex, accounting for its wide distribution at the leading edge.
利用光学显微镜和高分辨率电子显微镜,我们分析了在表皮生长因子刺激后,Arp2/3复合体与乳腺癌细胞片状伪足延伸所需的成核活性之间的时空关系。刺激后,丝状肌动蛋白的带刺端数量迅速短暂增加两到四倍,且几乎完全局限于延伸的片状伪足的最外边缘(质膜内100 - 200纳米范围内)。这伴随着前沿处丝状肌动蛋白密度的增加和丝状肌动蛋白长度的普遍缩短,尤其是长丝状肌动蛋白的减少。与此同时,Arp2/3复合体在整个皮质丝状肌动蛋白网络中被募集,其数量增加了1.5倍,该网络从前沿膜向深度延伸1 - 1.5微米。Arp2/3复合体在延伸的片状伪足膜处的募集表明,Arp2/3可能参与了生长中丝状肌动蛋白的初始生成。然而,皮质网络中存在的该复合体只有一小部分在游离带刺端附近共定位。这表明,在延伸的片状伪足前沿的100 - 200纳米的亚膜区构成了一个特殊的生化微环境,可能通过局部活跃的Arp2/3复合体、切断或降低封端蛋白的结合速率,有利于游离带刺端的生成和维持。我们的结果与解封是成核活性产生的主要机制这一假设不一致。然而,它们支持了Arp2/3介导的对通过切断等其他机制在膜附近形成的肌动蛋白寡聚体的捕获这一假设。它们还支持Arp2/3复合体对 pointed-end 的封端作用,这解释了其在前缘的广泛分布。