Hill E, van Der Kaay J, Downes C P, Smythe E
Division of Molecular Cell Biology, Wellcome Trust Biocentre, Dundee DD1 5EH, United Kingdom.
J Cell Biol. 2001 Jan 22;152(2):309-23. doi: 10.1083/jcb.152.2.309.
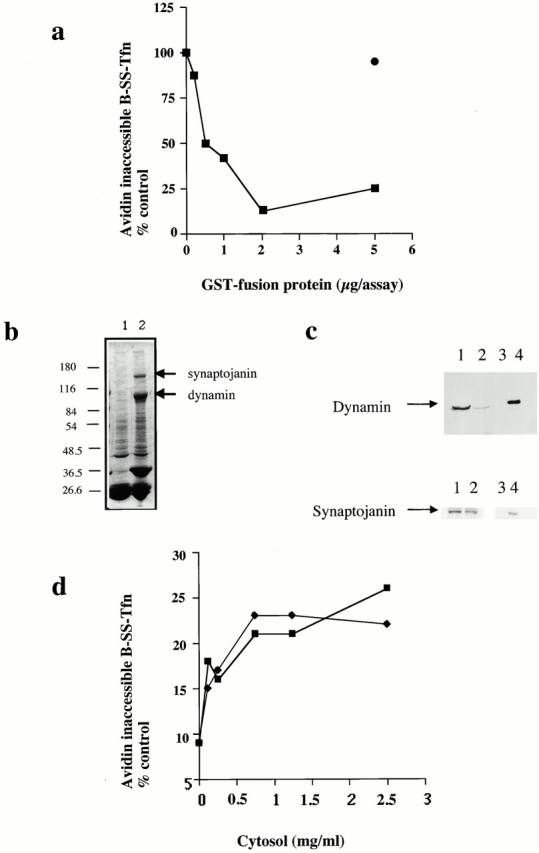
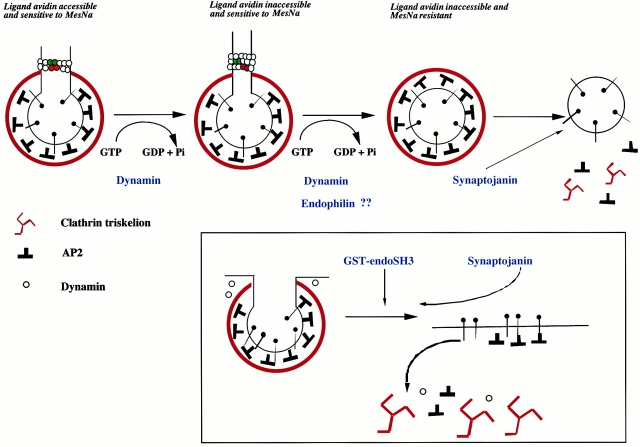
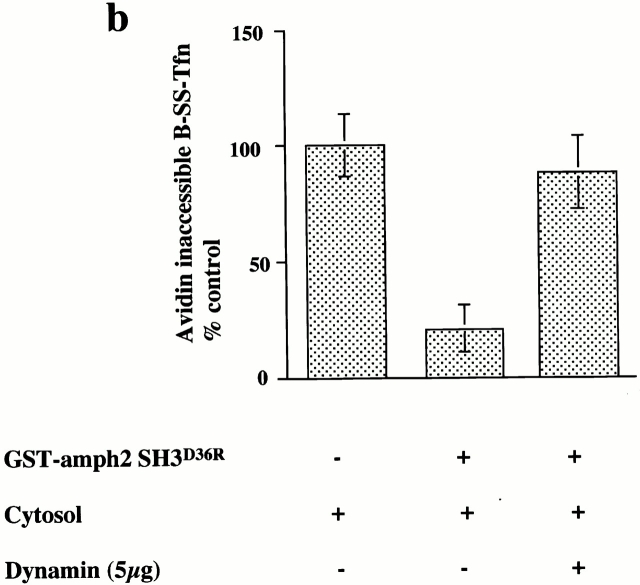
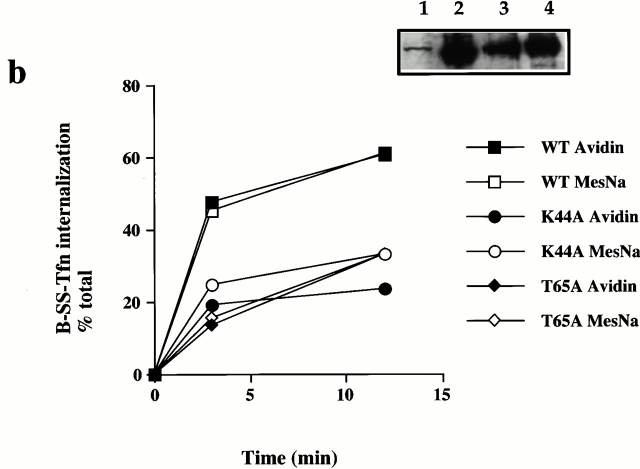
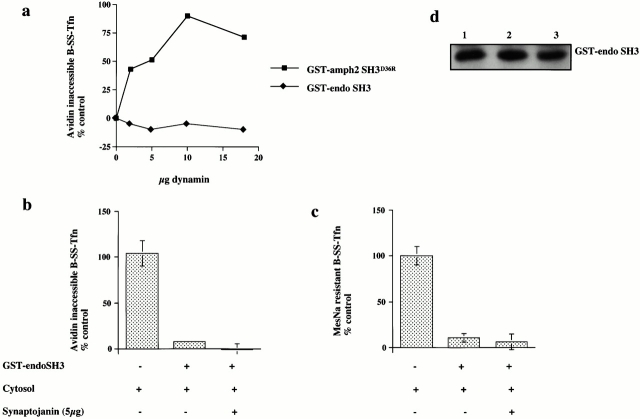
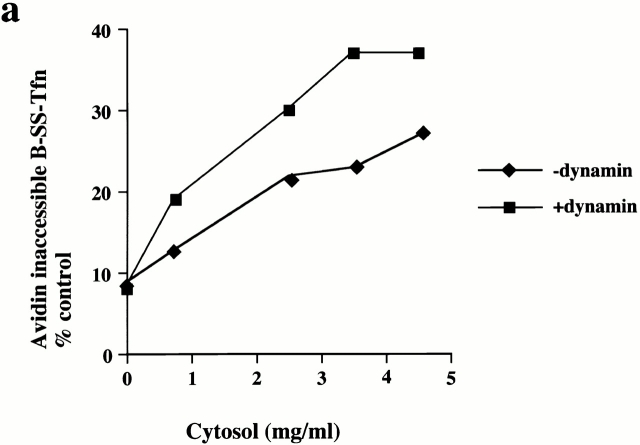
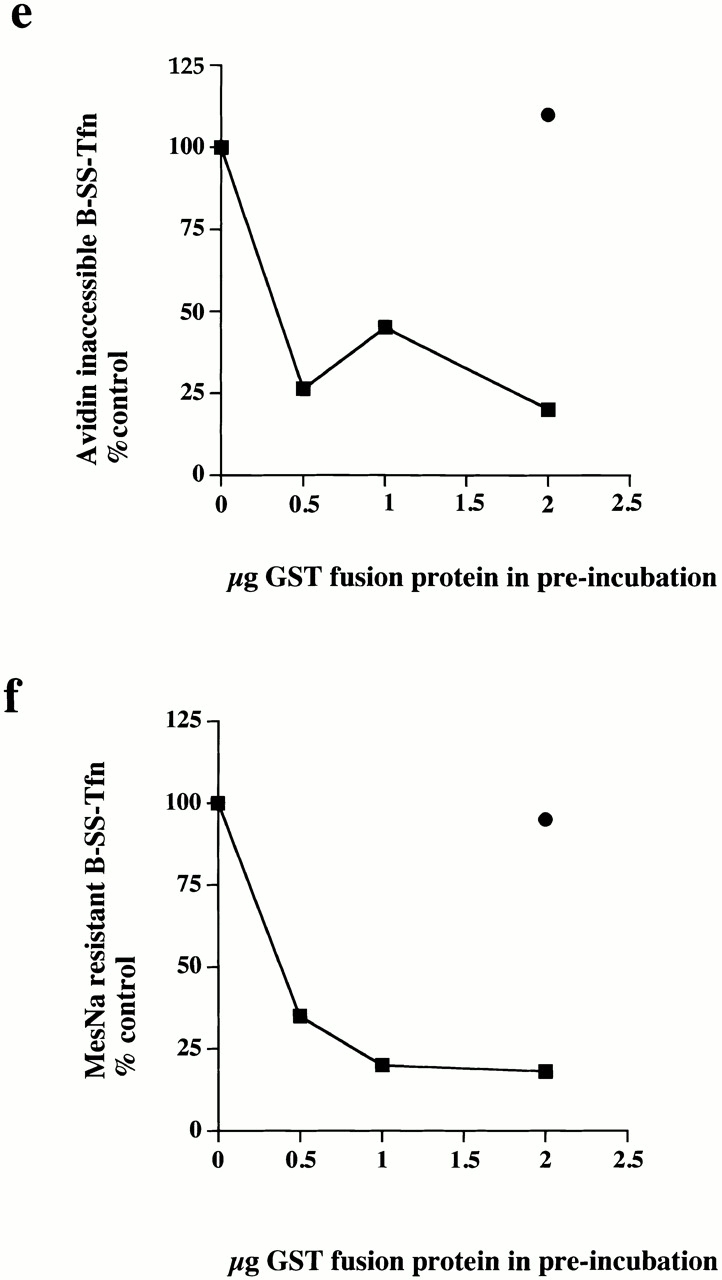
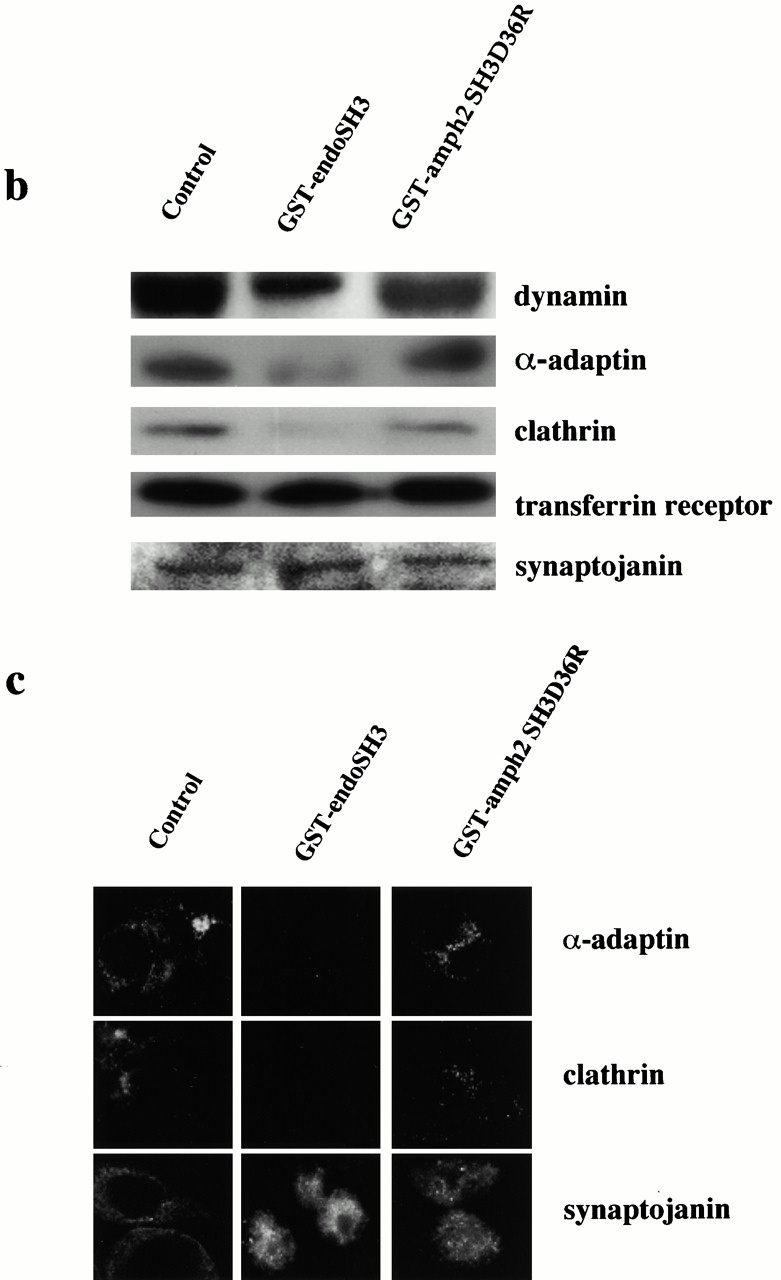
Plasma membrane clathrin-coated vesicles form after the directed assembly of clathrin and the adaptor complex, AP2, from the cytosol onto the membrane. In addition to these structural components, several other proteins have been implicated in clathrin-coated vesicle formation. These include the large molecular weight GTPase, dynamin, and several Src homology 3 (SH3) domain-containing proteins which bind to dynamin via interactions with its COOH-terminal proline/arginine-rich domain (PRD). To understand the mechanism of coated vesicle formation, it is essential to determine the hierarchy by which individual components are targeted to and act in coated pit assembly, invagination, and scission. To address the role of dynamin and its binding partners in the early stages of endocytosis, we have used well-established in vitro assays for the late stages of coated pit invagination and coated vesicle scission. Dynamin has previously been shown to have a role in scission of coated vesicles. We show that dynamin is also required for the late stages of invagination of clathrin-coated pits. Furthermore, dynamin must bind and hydrolyze GTP for its role in sequestering ligand into deeply invaginated coated pits. We also demonstrate that the SH3 domain of endophilin, which binds both synaptojanin and dynamin, inhibits both late stages of invagination and also scission in vitro. This inhibition results from a reduction in phosphoinositide 4,5-bisphosphate levels which causes dissociation of AP2, clathrin, and dynamin from the plasma membrane. The dramatic effects of the SH3 domain of endophilin led us to propose a model for the temporal order of addition of endophilin and its binding partner synaptojanin in the coated vesicle cycle.
网格蛋白包被小泡在网格蛋白和衔接蛋白复合体AP2从胞质溶胶定向组装到膜上之后形成。除了这些结构成分外,还有其他几种蛋白质也参与了网格蛋白包被小泡的形成。这些蛋白质包括大分子量的GTP酶发动蛋白,以及几种含Src同源结构域3(SH3)的蛋白质,它们通过与其COOH末端富含脯氨酸/精氨酸的结构域(PRD)相互作用而与发动蛋白结合。为了理解包被小泡形成的机制,确定各个成分靶向包被凹陷并在其中发挥作用、内陷和切割的层次结构至关重要。为了研究发动蛋白及其结合伙伴在内吞作用早期阶段的作用,我们使用了成熟的体外检测方法来研究包被凹陷内陷和包被小泡切割的后期阶段。先前已证明发动蛋白在包被小泡的切割中起作用。我们发现发动蛋白对于网格蛋白包被凹陷的后期内陷也是必需的。此外,发动蛋白必须结合并水解GTP才能在将配体隔离到深陷的包被凹陷中发挥作用。我们还证明,内吞蛋白的SH3结构域既能结合突触素I又能结合发动蛋白,它在体外抑制内陷后期阶段以及切割过程。这种抑制作用是由于4,5-二磷酸磷脂酰肌醇水平降低导致AP2、网格蛋白和发动蛋白从质膜上解离所致。内吞蛋白SH3结构域的显著作用使我们提出了一个关于内吞蛋白及其结合伙伴突触素I在包被小泡循环中添加的时间顺序的模型。