Mak D O, McBride S, Foskett J K
Department of Physiology, University of Pennsylvania, Philadelphia, Pennsylvania 19104, USA.
J Gen Physiol. 2001 Apr;117(4):299-314. doi: 10.1085/jgp.117.4.299.
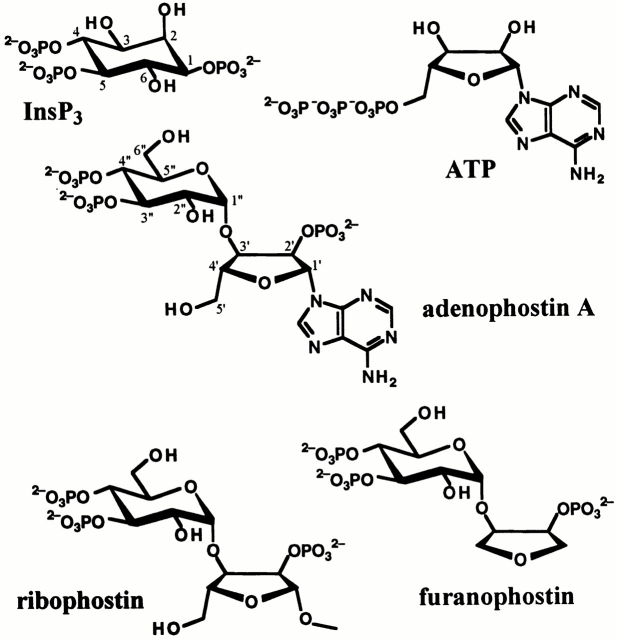
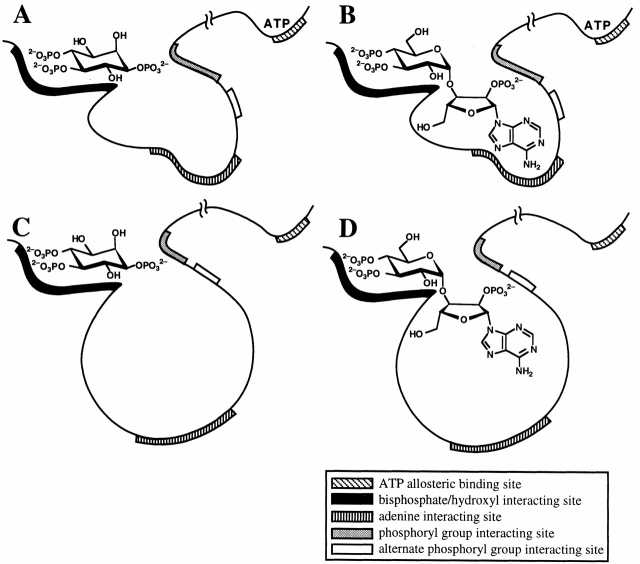
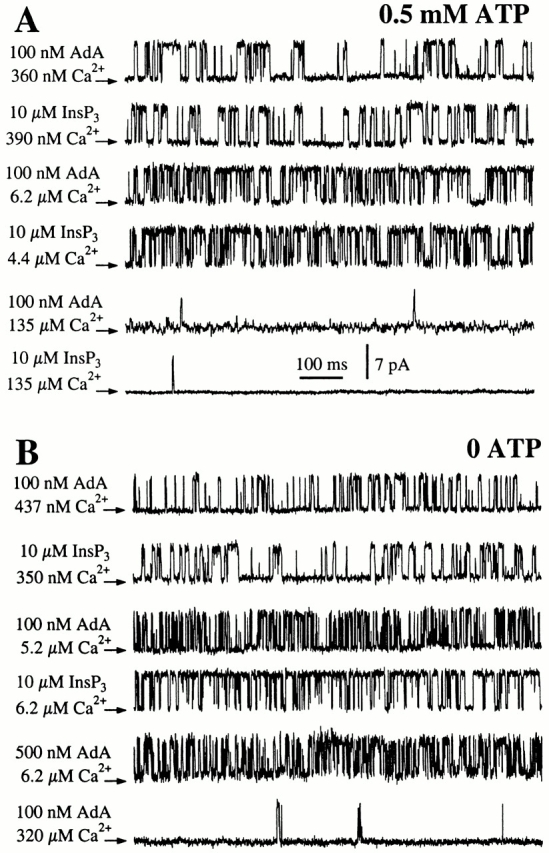
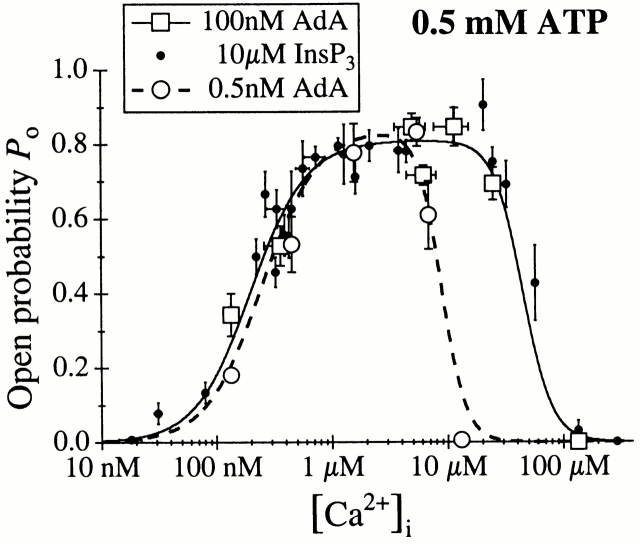
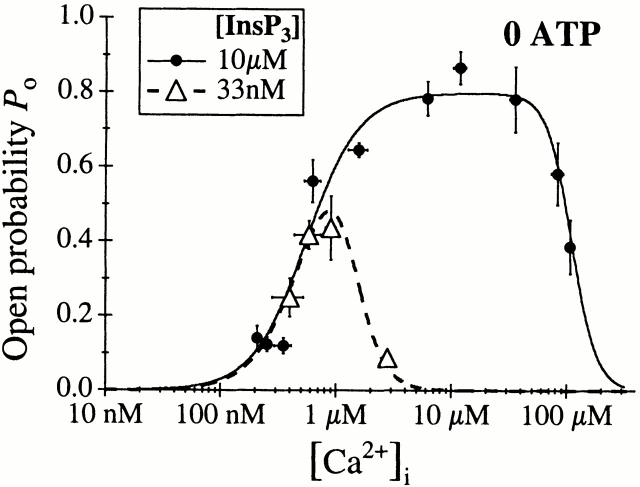
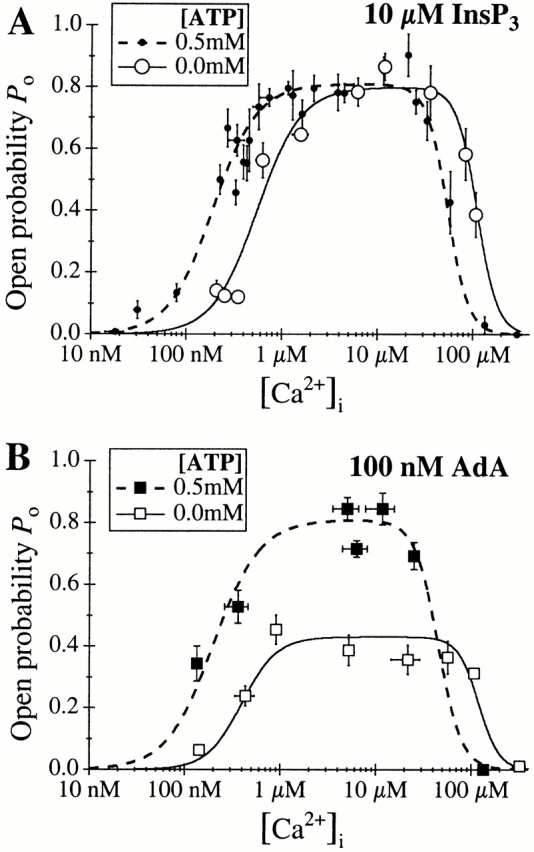
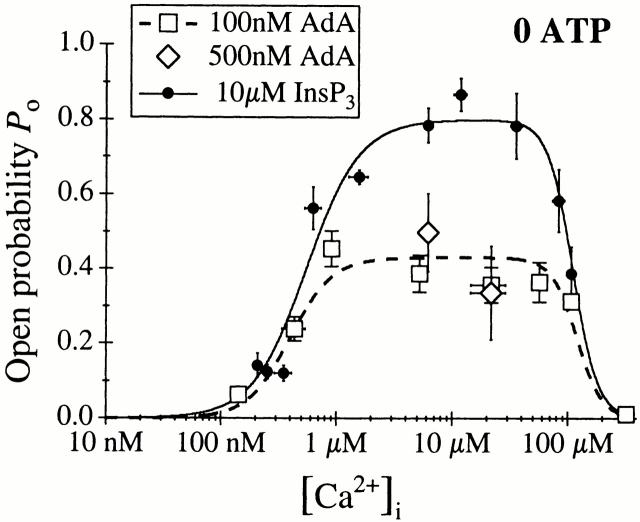
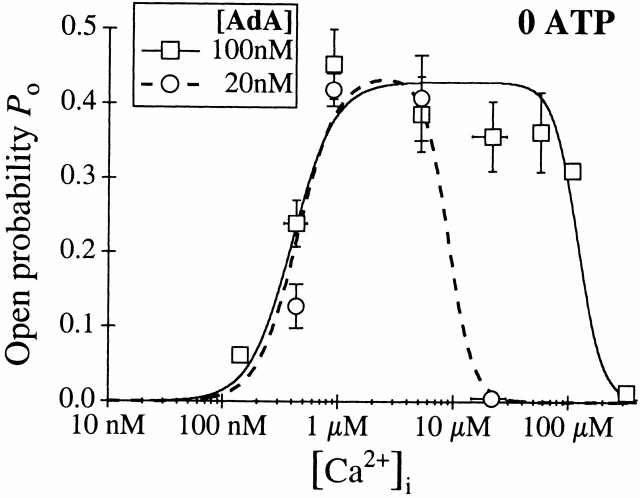
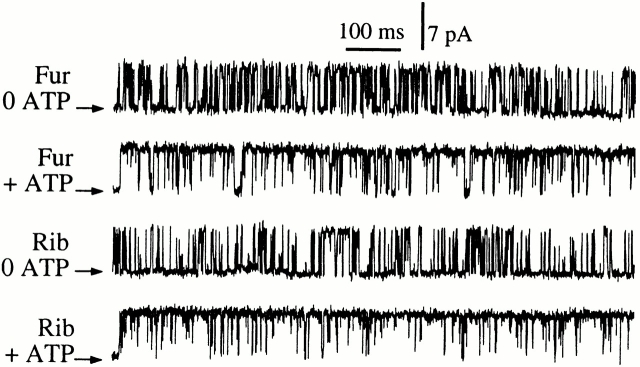
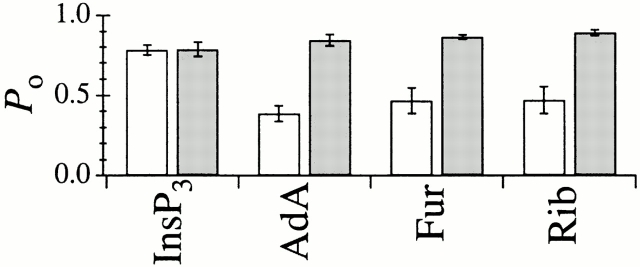
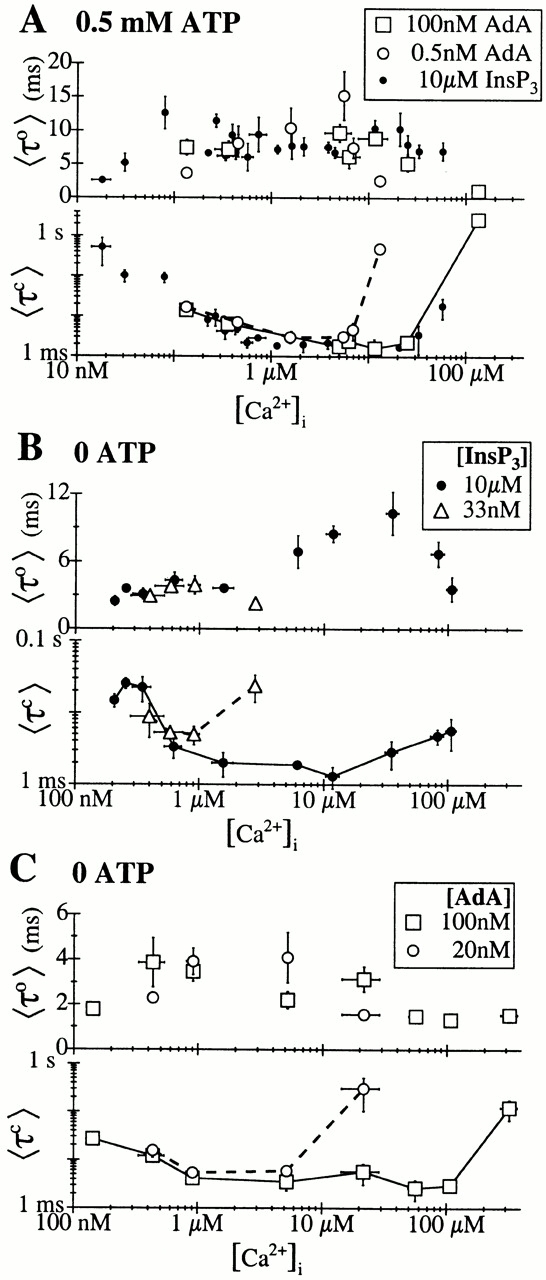
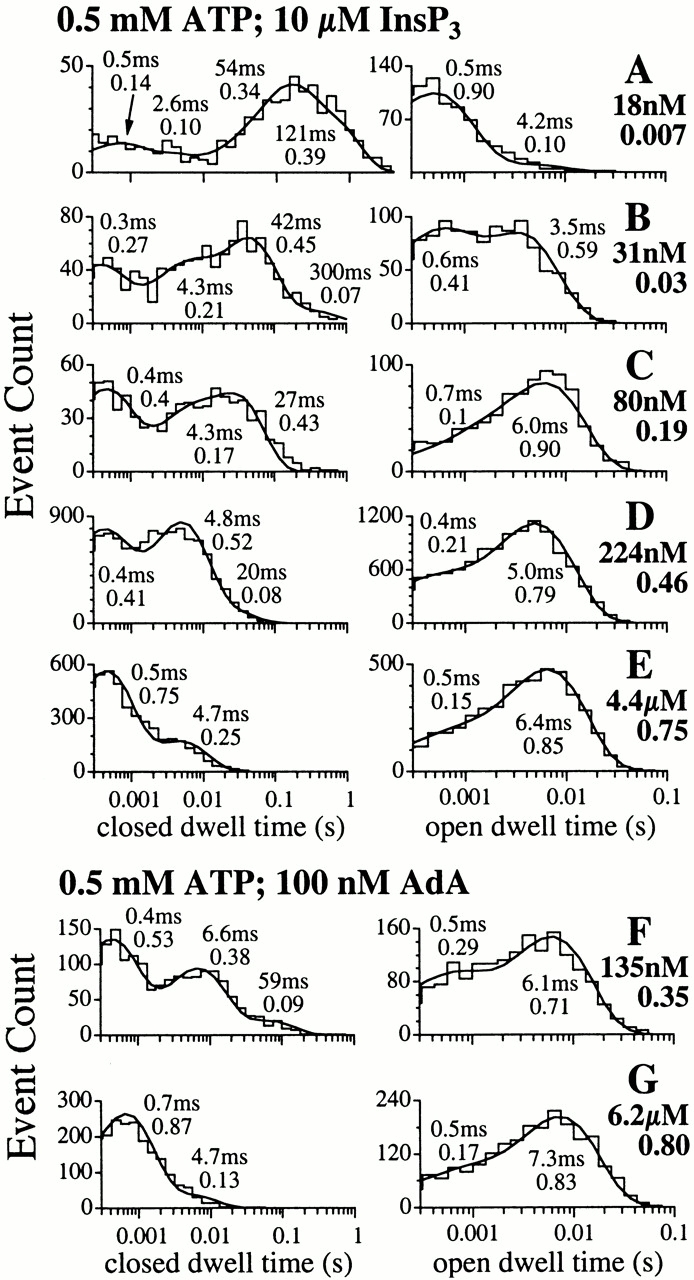
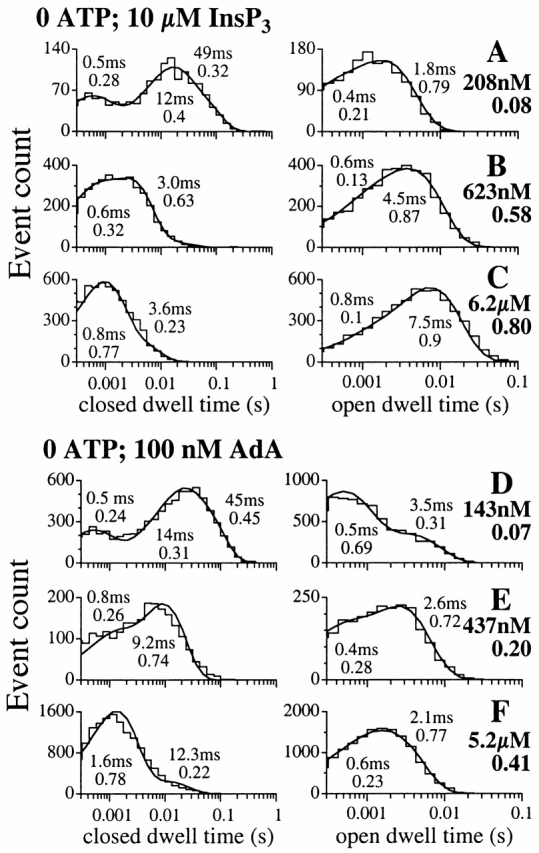
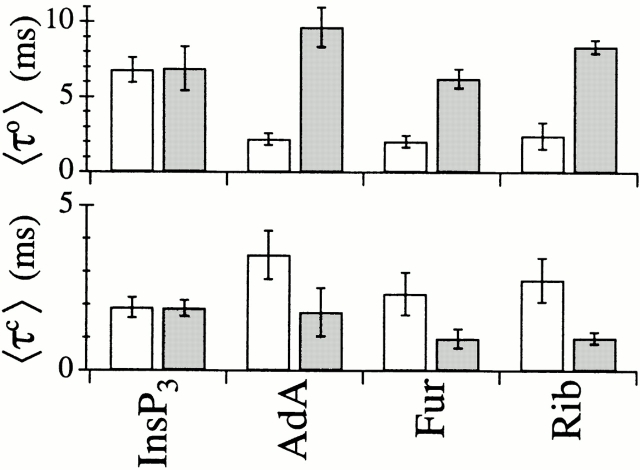
The inositol 1,4,5-trisphosphate (InsP(3)) receptor (InsP(3)R) is a ligand-gated intracellular Ca(2+) release channel that plays a central role in modulating cytoplasmic free Ca(2+) concentration (Ca(2+)). The fungal metabolite adenophostin A (AdA) is a potent agonist of the InsP(3)R that is structurally different from InsP(3) and elicits distinct calcium signals in cells. We have investigated the effects of AdA and its analogues on single-channel activities of the InsP(3)R in the outer membrane of isolated Xenopus laevis oocyte nuclei. InsP(3)R activated by either AdA or InsP(3) have identical channel conductance properties. Furthermore, AdA, like InsP(3), activates the channel by tuning Ca(2+) inhibition of gating. However, gating of the AdA-liganded InsP(3)R has a critical dependence on cytoplasmic ATP free acid concentration not observed for InsP(3)-liganded channels. Channel gating activated by AdA is indistinguishable from that elicited by InsP(3) in the presence of 0.5 mM ATP, although the functional affinity of the channel is 60-fold higher for AdA. However, in the absence of ATP, gating kinetics of AdA-liganded InsP(3)R were very different. Channel open time was reduced by 50%, resulting in substantially lower maximum open probability than channels activated by AdA in the presence of ATP, or by InsP(3) in the presence or absence of ATP. Also, the higher functional affinity of InsP(3)R for AdA than for InsP(3) is nearly abolished in the absence of ATP. Low affinity AdA analogues furanophostin and ribophostin activated InsP(3)R channels with gating properties similar to those of AdA. These results provide novel insights for interpretations of observed effects of AdA on calcium signaling, including the mechanisms that determine the durations of elementary Ca(2+) release events in cells. Comparisons of single-channel gating kinetics of the InsP(3)R activated by InsP(3), AdA, and its analogues also identify molecular elements in InsP(3)R ligands that contribute to binding and activation of channel gating.
肌醇1,4,5 -三磷酸(InsP(3))受体(InsP(3)R)是一种配体门控的细胞内Ca(2+)释放通道,在调节细胞质游离Ca(2+)浓度(Ca(2+))中起核心作用。真菌代谢产物腺嘌呤磷酯素A(AdA)是InsP(3)R的一种强效激动剂,其结构与InsP(3)不同,并在细胞中引发不同的钙信号。我们研究了AdA及其类似物对分离的非洲爪蟾卵母细胞核外膜中InsP(3)R单通道活性的影响。由AdA或InsP(3)激活的InsP(3)R具有相同的通道电导特性。此外,AdA与InsP(3)一样,通过调节Ca(2+)对门控的抑制来激活通道。然而,AdA配体化的InsP(3)R的门控对细胞质ATP游离酸浓度有至关重要的依赖性,而InsP(3)配体化的通道则未观察到这种依赖性。在存在0.5 mM ATP的情况下,由AdA激活的通道门控与由InsP(3)引发的门控无法区分,尽管通道对AdA的功能亲和力高60倍。然而,在没有ATP的情况下,AdA配体化的InsP(3)R的门控动力学非常不同。通道开放时间减少了50%,导致最大开放概率比在存在ATP时由AdA激活的通道,或在存在或不存在ATP时由InsP(3)激活的通道显著更低。此外,在没有ATP的情况下,InsP(3)R对AdA的功能亲和力高于对InsP(3)的功能亲和力几乎消失。低亲和力的AdA类似物呋喃磷酯素和核糖磷酯素激活InsP(3)R通道,其门控特性与AdA相似。这些结果为解释观察到的AdA对钙信号的影响提供了新的见解,包括决定细胞中基本Ca(2+)释放事件持续时间的机制。对由InsP(3)、AdA及其类似物激活的InsP(3)R单通道门控动力学的比较也确定了InsP(3)R配体中有助于通道门控结合和激活的分子元件。