D'Souza Lisa M, Zhong Jin
Institute for Cellular and Molecular Biology, Department of Chemistry and Biochemistry, and Section of Molecular Genetics and Microbiology, School of Biological Sciences, University of Texas at Austin, 78712, USA.
BMC Mol Biol. 2002 Dec 20;3:17. doi: 10.1186/1471-2199-3-17.
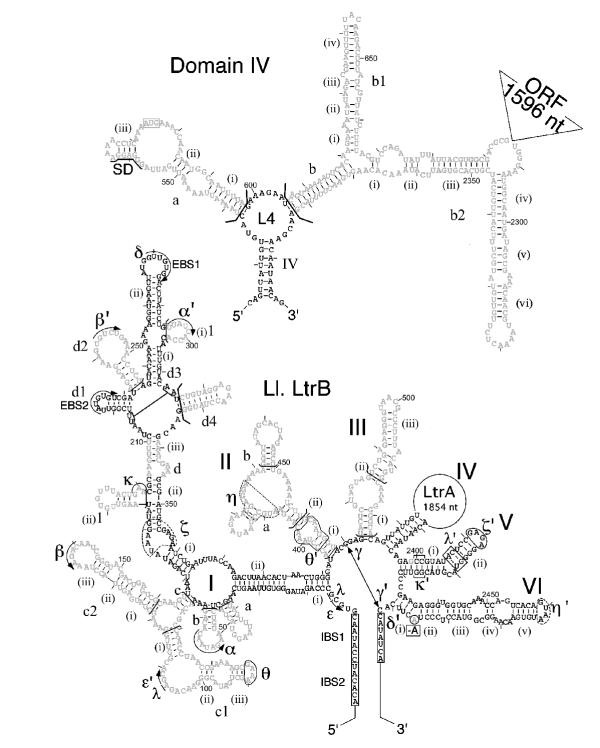
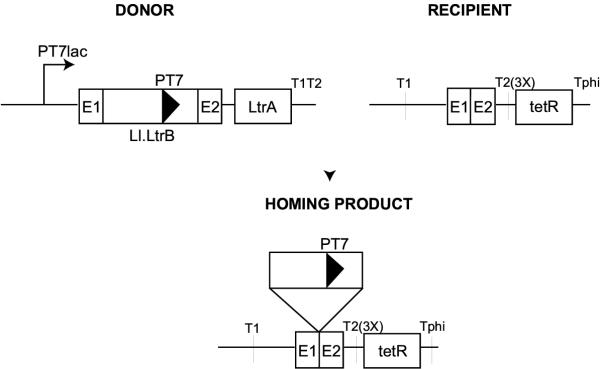
Group II introns are mobile genetic elements that form conserved secondary and tertiary structures. In order to determine which of the conserved structural elements are required for mobility, a series of domain and sub-domain deletions were made in the Lactococcus lactis group II intron (Ll.LtrB) and tested for mobility in a genetic assay. Point mutations in domains V and VI were also tested.
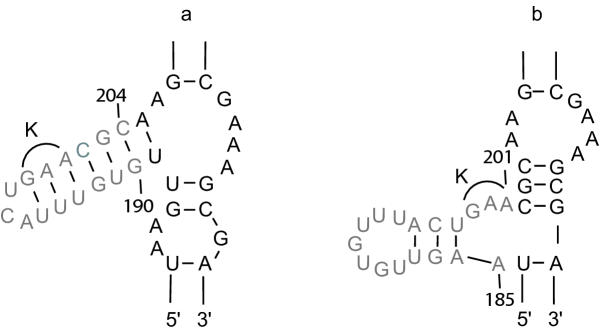
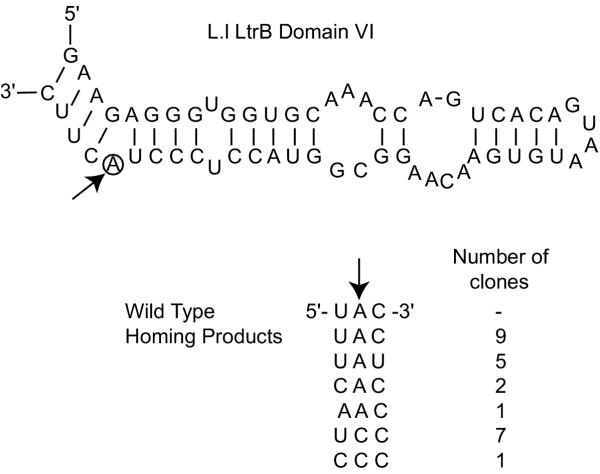
The largest deletion that could be made without severely compromising mobility was 158 nucleotides in DIVb(1-2). This mutant had a mobility frequency comparable to the wild-type Ll.LtrB intron (DeltaORF construct). Hence, all subsequent mutations were done in this mutant background. Deletion of DIIb reduced mobility to approximately 18% of wild-type, while another deletion in domain II (nts 404-459) was mobile to a minor extent. Only two deletions in DI and none in DIII were tolerated. Some mobility was also observed for a DIVa deletion mutant. Of the three point mutants at position G3 in DV, only G3A retained mobility. In DVI, deletion of the branch-point nucleotide abolished mobility, but the presence of any nucleotide at the branch-point position restored mobility to some extent.
The smallest intron capable of efficient retrohoming was 725 nucleotides, comprising the DIVb(1-2) and DII(ii)a,b deletions. The tertiary elements found to be nonessential for mobility were alpha, kappa and eta. In DV, only the G3A mutant was mobile. A branch-point residue is required for intron mobility.
II 组内含子是形成保守二级和三级结构的可移动遗传元件。为了确定哪些保守结构元件是移动所必需的,在乳酸乳球菌 II 组内含子(Ll.LtrB)中进行了一系列结构域和亚结构域缺失,并在遗传分析中测试其移动性。还测试了结构域 V 和 VI 中的点突变。
在不严重影响移动性的情况下可进行的最大缺失是 DIVb(1-2) 中的 158 个核苷酸。该突变体的移动频率与野生型 Ll.LtrB 内含子(DeltaORF 构建体)相当。因此,所有后续突变均在该突变体背景下进行。DIIb 的缺失将移动性降低至野生型的约 18%,而结构域 II 中的另一个缺失(核苷酸 404 - 459)移动性较小。DI 中只有两个缺失可被容忍,DIII 中则没有。DIVa 缺失突变体也观察到了一些移动性。在 DV 中 G3 位点的三个点突变体中,只有 G3A 保留了移动性(此处原文有误,应为“在 DV 中 G3 位点的三个点突变体中,只有 G3A 保留了移动性”)。在 DVI 中,分支点核苷酸的缺失消除了移动性,但在分支点位置存在任何核苷酸都能在一定程度上恢复移动性。
能够有效归巢的最小内含子为 725 个核苷酸,包括 DIVb(1-2) 和 DII(ii)a、b 缺失。发现对移动性非必需的三级元件是α、κ和η。在 DV 中,只有 G3A 突变体具有移动性。内含子移动需要一个分支点残基。