Shi Jiahai, Wei Zheng, Song Jianxing
Department of Biochemistry, National University of Singapore, 10 Kent Ridge Crescent, Singapore 119260.
J Biol Chem. 2004 Jun 4;279(23):24765-73. doi: 10.1074/jbc.M311744200. Epub 2004 Mar 22.
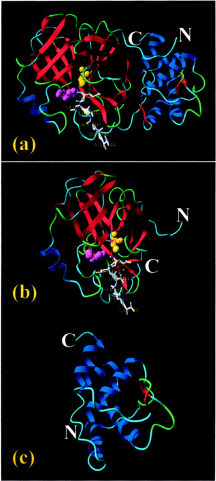
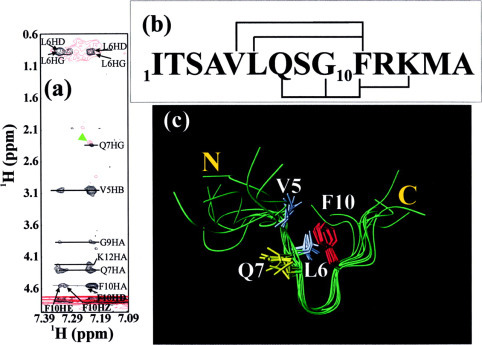
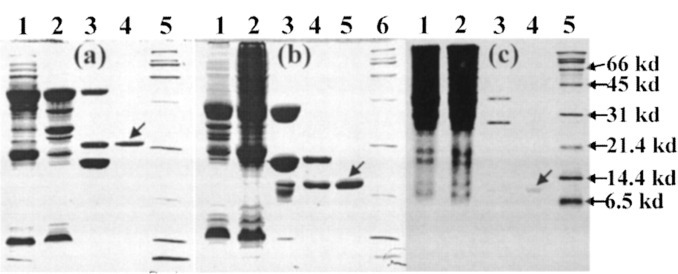
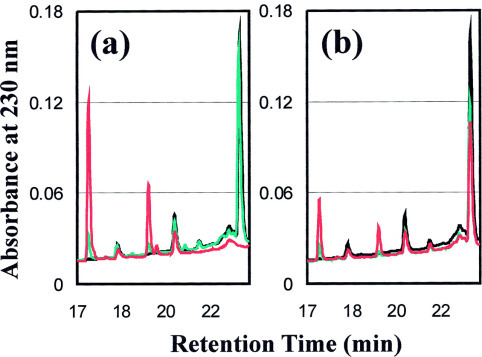
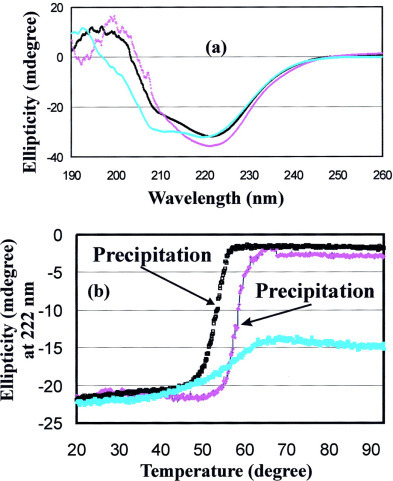
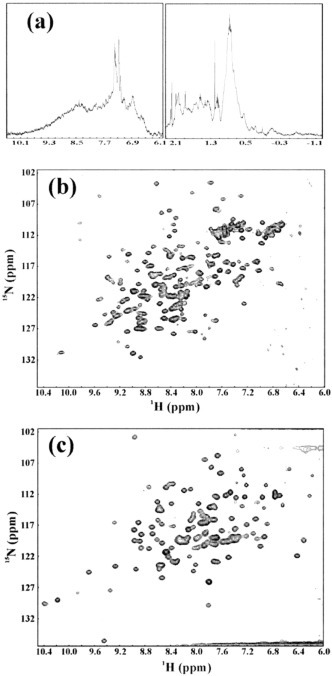
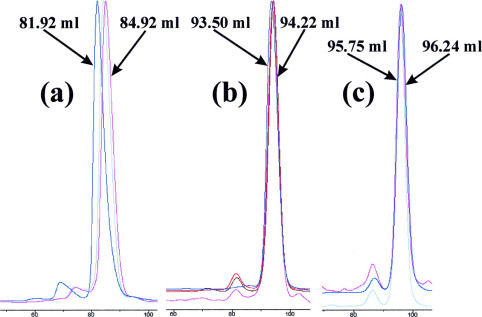
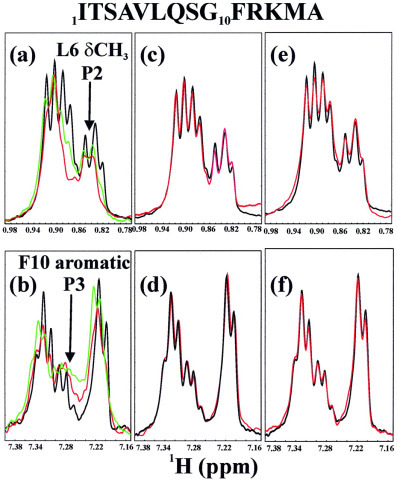
The severe acute respiratory syndrome (SARS) 3C-like protease consists of two distinct folds, namely the N-terminal chymotrypsin fold containing the domains I and II hosting the complete catalytic machinery and the C-terminal extra helical domain III unique for the coronavirus 3CL proteases. Previously the functional role of this extra domain has been completely unknown, and it was believed that the coronavirus 3CL proteases share the same enzymatic mechanism with picornavirus 3C proteases, which contain the chymotrypsin fold but have no extra domain. To understand the functional role of the extra domain and to characterize the enzyme-substrate interactions by use of the dynamic light scattering, circular dichroism, and NMR spectroscopy, we 1) dissected the full-length SARS 3CL protease into two distinct folds and subsequently investigated their structural and dimerization properties and 2) studied the structural and binding interactions of three substrate peptides with the entire enzyme and its two dissected folds. The results lead to several findings; 1) although two dissected parts folded into the native-like structures, the chymotrypsin fold only had weak activity as compared with the entire enzyme, and 2) although the chymotrypsin fold remained a monomer within a wide range of protein concentrations, the extra domain existed as a stable dimer even at a very low concentration. This observation strongly indicates that the extra domain contributes to the dimerization of the SARS 3CL protease, thus, switching the enzyme from the inactive form (monomer) to the active form (dimer). This discovery not only separates the coronavirus 3CL protease from the picornavirus 3C protease in terms of the enzymatic mechanism but also defines the dimerization interface on the extra helical domain as a new target for design of the specific protease inhibitors. Furthermore, the determination of the preferred solution conformation of the substrate peptide S1 together with the NMR differential line-broadening and transferred nuclear Overhauser enhancement study allows us to pinpoint the bound structure of the S1 peptide.
严重急性呼吸综合征(SARS)3C样蛋白酶由两个不同的折叠结构组成,即N端胰凝乳蛋白酶折叠结构,包含负责完整催化机制的结构域I和II,以及冠状病毒3CL蛋白酶特有的C端额外螺旋结构域III。此前,这个额外结构域的功能作用完全未知,人们认为冠状病毒3CL蛋白酶与小RNA病毒3C蛋白酶具有相同的酶促机制,后者含有胰凝乳蛋白酶折叠结构,但没有额外结构域。为了了解这个额外结构域的功能作用,并利用动态光散射、圆二色性和核磁共振光谱来表征酶-底物相互作用,我们进行了以下研究:1)将全长SARS 3CL蛋白酶分解为两个不同的折叠结构,随后研究它们的结构和二聚化特性;2)研究三种底物肽与完整酶及其两个分解后的折叠结构的结构和结合相互作用。研究结果有以下几点:1)尽管两个分解后的部分折叠成了类似天然的结构,但与完整酶相比,胰凝乳蛋白酶折叠结构的活性较弱;2)尽管胰凝乳蛋白酶折叠结构在很宽的蛋白质浓度范围内都保持单体状态,但额外结构域即使在非常低的浓度下也以稳定的二聚体形式存在。这一观察结果有力地表明,额外结构域有助于SARS 3CL蛋白酶的二聚化,从而将酶从无活性形式(单体)转变为活性形式(二聚体)。这一发现不仅在酶促机制方面将冠状病毒3CL蛋白酶与小RNA病毒3C蛋白酶区分开来,还将额外螺旋结构域上的二聚化界面定义为设计特异性蛋白酶抑制剂的新靶点。此外,通过确定底物肽S1的优选溶液构象,以及核磁共振差分线宽展宽和转移核Overhauser增强研究,我们能够确定S1肽的结合结构。