Wang Guangyu, Shahidullah Mohammad, Rocha Carmen A, Strang Candace, Pfaffinger Paul J, Covarrubias Manuel
Department of Pathology, Anatomy, and Cell Biology, Jefferson Medical College of thomas Jefferson University, Philadelphia, PA 19107, USA.
J Gen Physiol. 2005 Jul;126(1):55-69. doi: 10.1085/jgp.200509288. Epub 2005 Jun 13.
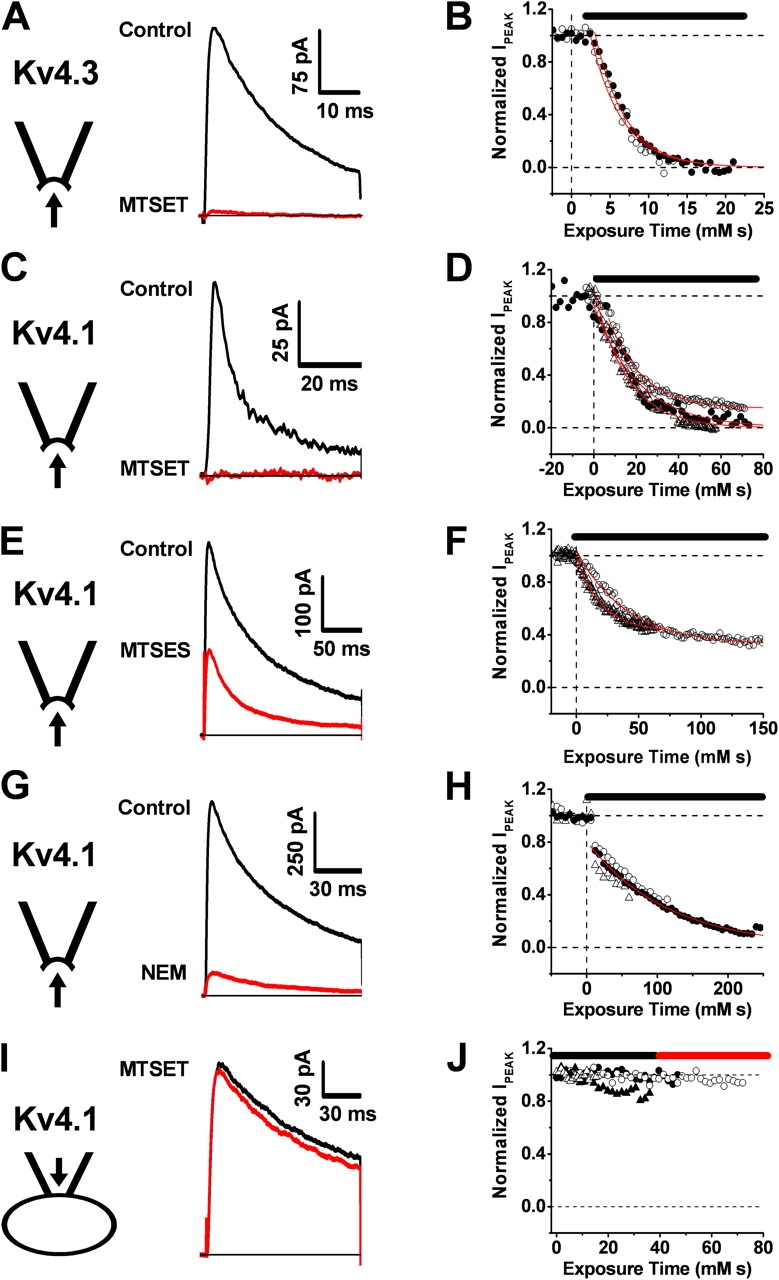
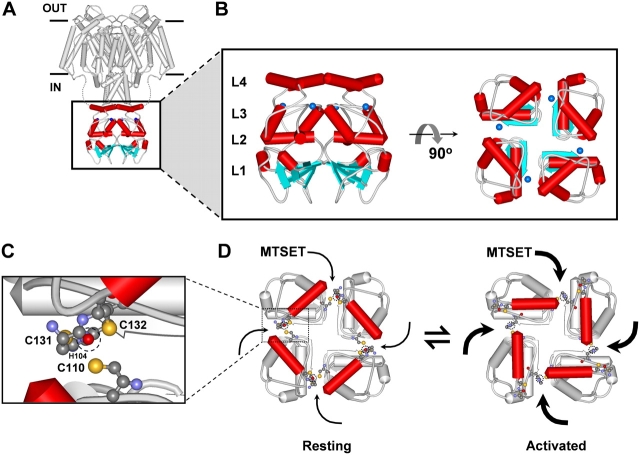
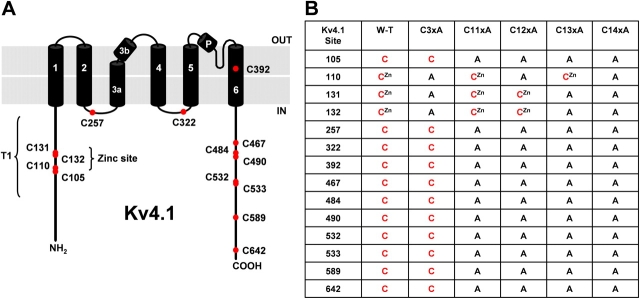
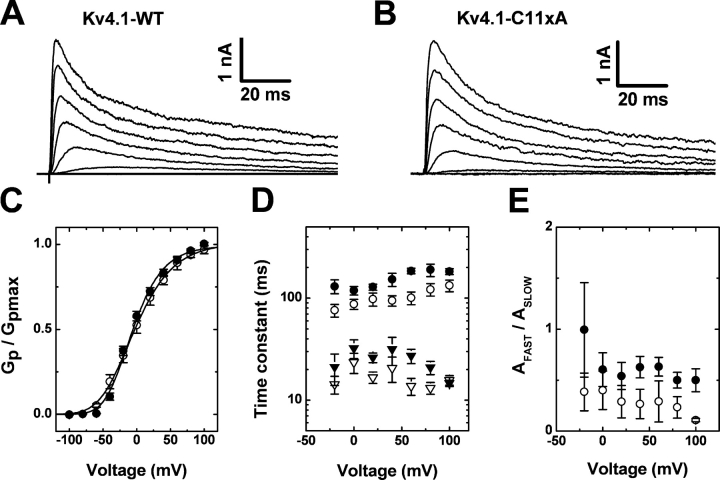
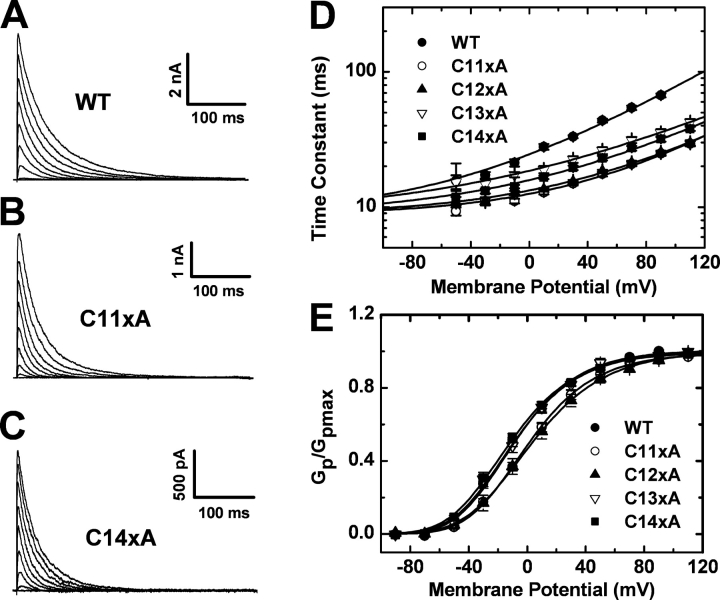
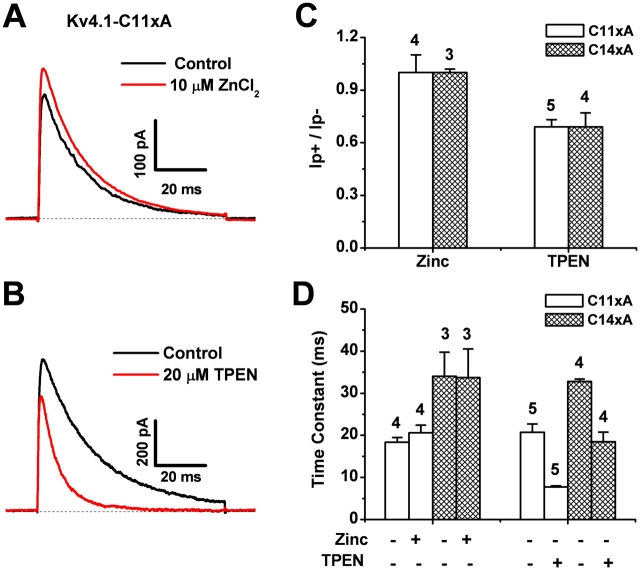
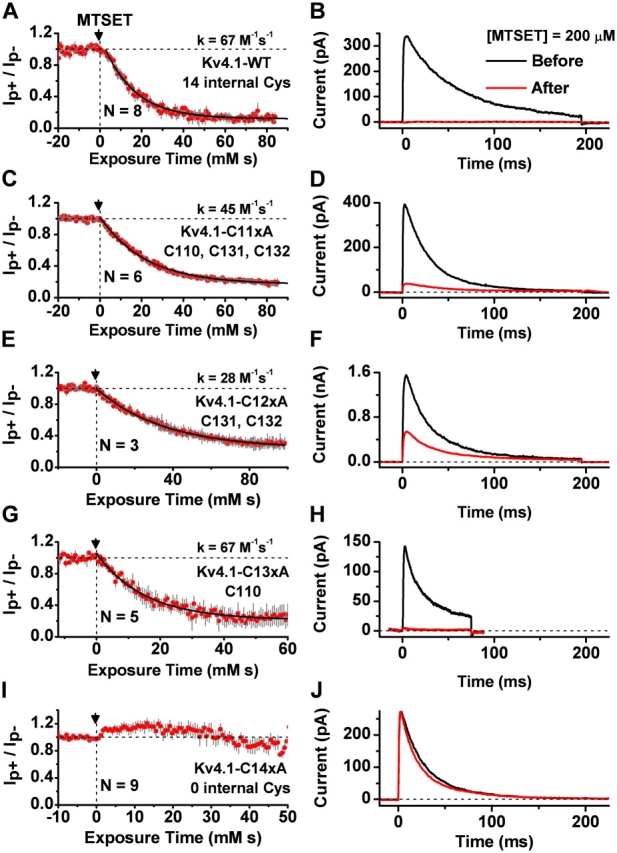
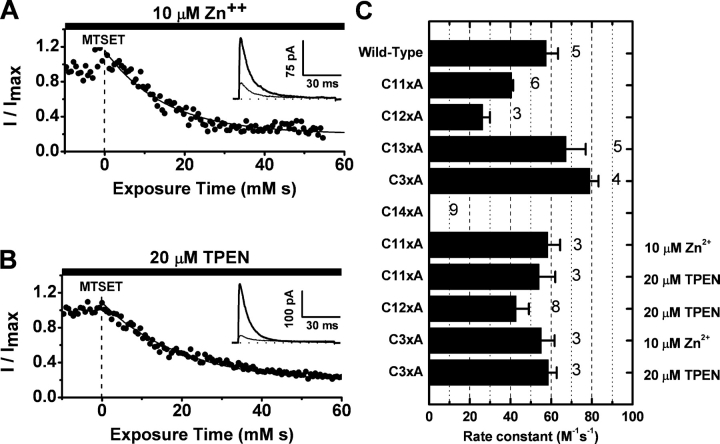
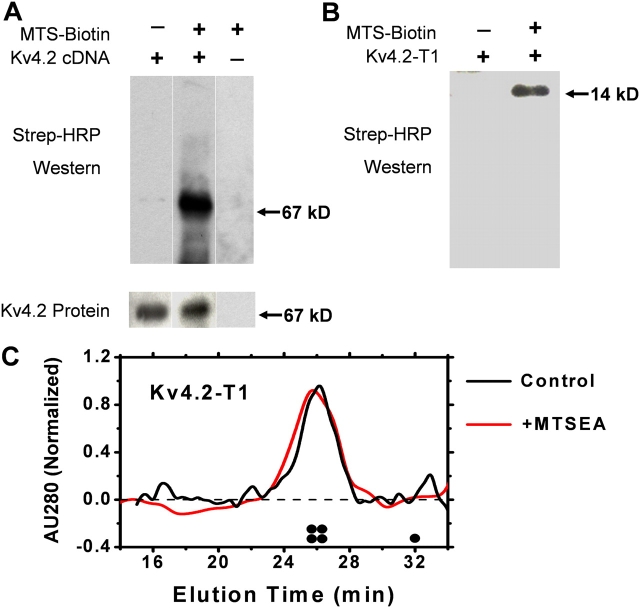
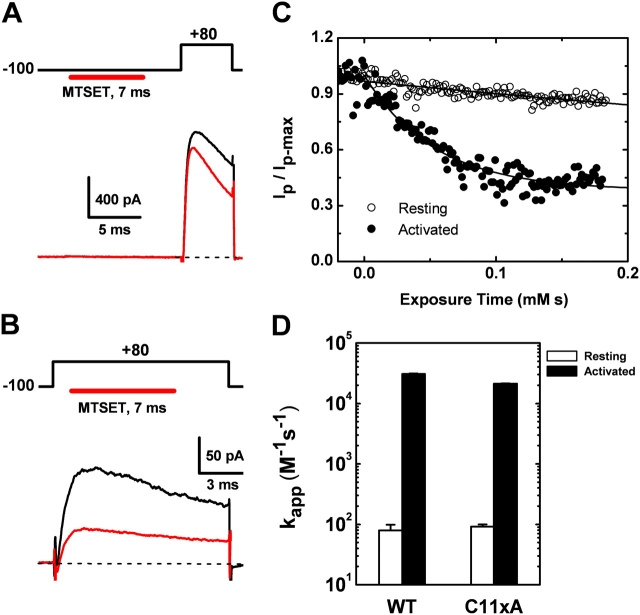
Gating of voltage-dependent K(+) channels involves movements of membrane-spanning regions that control the opening of the pore. Much less is known, however, about the contributions of large intracellular channel domains to the conformational changes that underlie gating. Here, we investigated the functional role of intracellular regions in Kv4 channels by probing relevant cysteines with thiol-specific reagents. We find that reagent application to the intracellular side of inside-out patches results in time-dependent irreversible inhibition of Kv4.1 and Kv4.3 currents. In the absence or presence of Kv4-specific auxiliary subunits, mutational and electrophysiological analyses showed that none of the 14 intracellular cysteines is essential for channel gating. C110, C131, and C132 in the intersubunit interface of the tetramerization domain (T1) are targets responsible for the irreversible inhibition by a methanethiosulfonate derivative (MTSET). This result is surprising because structural studies of Kv4-T1 crystals predicted protection of the targeted thiolate groups by constitutive high-affinity Zn(2+) coordination. Also, added Zn(2+) or a potent Zn(2+) chelator (TPEN) does not significantly modulate the accessibility of MTSET to C110, C131, or C132; and furthermore, when the three critical cysteines remained as possible targets, the MTSET modification rate of the activated state is approximately 200-fold faster than that of the resting state. Biochemical experiments confirmed the chemical modification of the intact alpha-subunit and the purified tetrameric T1 domain by MTS reagents. These results conclusively demonstrate that the T1--T1 interface of Kv4 channels is functionally active and dynamic, and that critical reactive thiolate groups in this interface may not be protected by Zn(2+) binding.
电压依赖性钾离子通道的门控涉及跨膜区域的运动,这些运动控制着通道孔的开放。然而,关于大的细胞内通道结构域对门控基础构象变化的贡献,我们所知甚少。在这里,我们通过用硫醇特异性试剂探测相关半胱氨酸,研究了Kv4通道中细胞内区域的功能作用。我们发现,将试剂应用于内向外膜片的细胞内侧会导致Kv4.1和Kv4.3电流随时间不可逆抑制。在不存在或存在Kv4特异性辅助亚基的情况下,突变和电生理分析表明,14个细胞内半胱氨酸中没有一个对通道门控至关重要。四聚化结构域(T1)亚基间界面中的C110、C131和C132是甲硫基磺酸盐衍生物(MTSET)不可逆抑制的靶点。这一结果令人惊讶,因为Kv4-T1晶体的结构研究预测,靶向硫醇盐基团会通过组成性高亲和力锌离子配位得到保护。此外,添加锌离子或强效锌离子螯合剂(TPEN)不会显著调节MTSET对C110、C131或C132的可及性;而且,当这三个关键半胱氨酸仍为可能的靶点时,激活状态下MTSET的修饰速率比静息状态快约200倍。生化实验证实了MTS试剂对完整α亚基和纯化的四聚体T1结构域的化学修饰。这些结果确凿地证明,Kv4通道的T1-T1界面具有功能活性且是动态的,并且该界面中的关键反应性硫醇盐基团可能不受锌离子结合的保护。